Indledning
Smågrise dødelighed er et af de største udvalg træk inden for svineavl og er påvirket af soen, smågrise og miljø. Derfor er pigletdødelighed en kompleks fænotype og afhænger af soens kapacitet til at hæve sit afkom, men er også en funktion af fødselsvægt, ledelse og udvælgelse (Knol et al., 2002)., Imidlertid bidrager også monogene recessive defekter til pigletdødelighed, skønt der kun er rapporteret få eksempler tidligere (Murgiano et al., 2012; Matika et al., 2019). Selv i de tilfælde, hvor virkningen af mutationen er alvorlig, er valg effektivt mod en sådan mutation hæmmet af lavfrekvensen. I mange alvorlige defekter dør dieygoter meget tidligt i svangerskabet og efterlader intet andet spor end fraværet af Homo .ygoter i befolkningen som helhed (Derks et al., 2019).,
Indavlseffekter i kommercielle svinepopulationer holdes normalt i kontrol ved selektiv avl for nedsat dødelighed hos smågrise ved at forbedre både moderskabsevner og grise levedygtighed (Olijslagers, 2018). Imidlertid er varianter, der ligger til grund for recessive monogene defekter, ikke godt fanget inden for avlsværdierne og potentielt glider til højere frekvenser som et resultat af intens udvælgelse (Georges et al., 2019). Desuden kan disse varianter også opretholdes som et resultat af balancering udvælgelse for en korreleret positiv effekt i Hetero .ygot tilstand (Derks et al., 2018).,
Recessive defekter bidrager kun marginalt til den samlede pigletdødelighed (Alonso-Spilsbury et al., 2007). Ikke desto mindre er varianter, der påvirker svinedødeligheden, af stor betydning, fordi disse varianter direkte påvirker produktion og dyrevelfærd (Ba .ter et al., 2013; Rutherford et al., 2013). I dyrepopulationsstyring er lavfrekvent forekomst af defekter imidlertid normalt dårligt dokumenteret (ofte bruges meget generelle udtryk), og syndromer genkendes ofte kun, når de har nået en høj frekvens., Dette er især relevant for syndromer, der ikke fører til meget forskellige fænotyper. Selv i kommercielle avlspopulationer kan der derfor kun foretages lidt sporing af specifikke syndromer, og for effektivt at vælge mod specifikke lavfrekvente syndromer kræver derfor nye tilgange.
i dette arbejde beskriver vi opdagelsen af et stærkt svækkende syndrom i en kommerciel svinepopulation gennem en undersøgelse baseret på en kombineret SNP-arrays med medium densitet og sekventering af hele genomet (.gs)., Undersøgelsen førte til identifikation af en 16-bp frameshift-deletion i SPTBN4-genet med forudsagte klare fænotypiske konsekvenser i homo .ygoter. 9% i den undersøgte befolkning, hvilket påvirker cirka 0, 81% af befolkningens kuld. Frekvensen var tilstrækkelig lav til at være ukendt til at have et genetisk grundlag og endda effektivt blive ukendt som et specifikt syndrom overhovedet. Efter gennemførelsen af undersøgelsen blev en gravid sår identificeret af en bæresvin., De berørte smågrise lider af myopati og er ikke i stand til at gå, hvilket normalt resulterer i død inden for få timer efter fødslen, helt i overensstemmelse med forudsagt patologi i sammenligning med lignende menneskelige og mus tilfælde.
Materiale og Metoder
Dyr, Genotyper, Pre-Processing
datasættet består af 31,839 dyr fra en syntetisk vildsvin, på linje med store hvide baggrund. Linjen vedligeholdes og opdrættes i Topigs Norsvin nucleus gårde, primært udvælgelse på produktion og sundhed træk., Dyrene blev genotypet på Illumina GeneSeek custom 50K SNP chip (Lincoln, NE, USA). Dyr med en hyppighed af manglende genotyper > 0, 15 blev fjernet. Vi kasserede markører, der ikke opfyldte følgende filtreringskriterier: en minimumsopkaldshastighed på 0.85, en mindre allelfrekvens > 0.01, og en Hardy-.einberg-proportioner nøjagtig test p-værdi under P < 10-12. Desuden blev markører med ukendt placering på sscrofa11.1-genombygningen kasseret, hvilket efterlod 41,573 markører efter filtrering., Alle trin blev udført i Plink v1.90b3.30 (Purcell et al., 2007).
Haplotype Indfasning og Identifikation af SSC6 Haplotype
Vi har udført haplotype afvikling og beregning af mangler steder i Beagle5.0 med parameter for effektiv populationsstørrelse er sat til 100, andre indstillinger var standardindstillinger (Andersen et al., 2018). Forventede homo .ygoter blev estimeret baseret på haplotypefrekvens ved hjælp af Hardy-.einberg-princippet. En nøjagtig binomial test blev anvendt til at teste antallet af observerede homo .ygoter med antallet af forventede homo .ygoter., Haplotypen blev betragtet som signifikant udtømt, hvis P < 5 10 10-3.
Fænotypiske Effekter, der er Forbundet Med SSC6 Haplotype
Vi har undersøgt SSC6 haplotype for registreringer på det samlede antal fødte, antal dødfødte, mumificerede smågrise, fare overlevelse, og amning overlevelse (overlevelse op til omkring 21 dage gamle) i alt 9,666 kuld. Vi opført disse fænotyper for alle C .c, og C .n kuld identificeret. Vi brugte en telchs t-test til at vurdere, om fænotyperne fra C .c-kuldene adskiller sig markant fra C .n-kuldene. En p-værdi < 0.,05 blev betragtet som signifikant.
Hel-Genom-Sekventering Analyse og Kandidat Variant Identifikation
datasættet består af 71 hele genom sekventeret personer fra befolkningen under undersøgelse. Alle 71 prøver var også til stede i vores datasæt af 31,839 dyr genotyped på 50K. Den 71-prøver har et samlet volumen på 1.93 Tbp (tera-basepar) fra 14.16 milliarder 150-bp parret-end-læser (Tabel S3). Prøverne blev sekventeret på Illumina Hise.2000. Vi justerede sekvenserne til sscrofa11. 1 genombygningen ved hjælp af B .a-MEM version 0.7.,15 (Li og Durbin, 2009) med en gennemsnitlig mappability på 98.9% og en prøve dækning spænder fra 8.8 til 14.8. (10.9 average gennemsnit). Samblaster blev brugt til at fjerne PCR-duplikater (Faust and Hall, 2014). Samtools blev brugt til at sortere, flette og indeksere bam-filer (Li et al., 2009). Kortlægning og kvalitetsstatistikker blev genereret ved hjælp af Qualualimap (Okonechnikov et al., 2016). Variant kalde blev udført med Freebayes v1.1.0 med følgende indstillinger: –min-base-kvalitet 10 –min-alternativ-fraktion 0.2 –haplotype-længde 0 –min-alternativ-tæller 2 (Garrison og Marth, 2012)., Varianter med Phred quality score < 20 blev kasseret (Li et al., 2009). Varianter blev annoteret under anvendelse af Ensembl-varianteffektprediktoren (VEP, release 96) (Mclaren et al., 2016). Virkningen af missense-varianter blev forudsagt ved hjælp af sortering intolerant fra tolerant (SIFT) (Kumar et al., 2009). LD-analyse blev udført under anvendelse af Plink v1.90b3.30 (Purcell et al., 2007) med følgende indstillinger –chr-sæt 18, –r2, ld-vindue-r2 0.8.
SPTBN4 Proteinjustering
Proteinjustering mellem vildtype og mutant protein blev udført under anvendelse af ClustalO (Madeira et al.,, 2019) og visualiseret ved hjælp af ESPript 3 (Robert and Gouet, 2014). Yderligere visualisering og validering blev udført ved hjælp af jbro .se genome vie .er version 1.12.1 (Skinner et al., 2009).
Validering af Kausale 16 bp SPTBN4 Sletning
PCR blev gjort ved hjælp af 60 ng af det genomiske DNA, med 0,4 µm af hver primer, 1,8 mM MgCl2, og 25 enheder/ml OneTaq® DNA-Polymerase (OneTaq® 2X Master Mix med Standard Buffer, New England Biolabs) i producentens PCR buffer i en endelig mængde af 12 µl., Indledende denaturering i 1 min. ved 95°C blev efterfulgt af 35 cyklusser af 95°C til 30 s, 55°C til 45 s, 72°C 90 s, efterfulgt af en 5 min extension 72°C. PCR-primere til SPTBN4 er TCAAGGGTGCAGGCTCTTTC frem og GGTAGGAAGCTCGAAGTGGG omvendt. Den forreste primer blev farvemærket med enten 6-FAM for at fremstille et fluorescerende mærket PCR-produkt, der kunne påvises på ABI 3730 DNA-se .uencer (Applied Biosystems). Fragment størrelser blev bestemt ved hjælp af GeneMapper soft .are 5 fra ABI.,
histopatologisk undersøgelse
to berørte smågrise under 1 uge blev sendt til patologiafdelingen for Royal Animal Health (Deventer) til undersøgelse. Makroskopisk var alle observationer inden for normale grænser. Der blev udtaget prøver af skeletmuskulaturen i forbenet, rygmuskulaturen og bagbenet på begge dyr til rutinemæssig h&e-farvning og Ptah-farvning. Muskelvævet blev opbevaret i separate krukker og fikseret i formaldehydopløsning 4%, bufret (=formalinopløsning 10%, bufret)., Derefter blev vævet indlejret i paraffin og skåret i 2 µm i henhold til standardoperationsprocedure (SOP RAH). Derefter blev objektglassene afparaffiniseret og rutinemæssigt farvet for hæmato .ylin og eosin (H&E) i en automatisk farvemaskine. Samtidig blev der fremstillet yderligere lysbilleder af 2 um af muskelvævet samt et positivt kontrolglide af muskelvæv til manuel farvning med “phosphotungstic acid hemato .ylin”, forkortet som PTAH. Denne farvning foretrækkes til at demonstrere tværstrimler af skeletmuskulatur.,
avlsværdier og Foreningsanalyse
i denne undersøgelse vurderede vi 63 træk, der blev brugt i avlsprogrammet. Deregulerede estimerede avlsværdier (DEBV) blev brugt som en responsvariabel for hvert træk under undersøgelse. Den estimerede avlsværdi (EBV) af alle evaluerede træk blev afregistreret ved hjælp af den metode, der er beskrevet af Garrick et al. (2009). EBV for hvert dyr blev opnået fra den rutinemæssige genetiske evaluering ved et kommercielt avlsprogram (Topigs Norsvin) ved hjælp af en dyremodel., Dyr med henblik på deregression blev ekstraheret fra den genetiske evaluering baseret på metodologien for Tier og Meyer (2004). De arveligheder, der blev brugt til deregression, blev også ekstraheret fra den rutinemæssige genetiske evaluering. Endelig blev vægtningsfaktorer baseret på den estimerede pålidelighed af DEBV også estimeret ifølge Garrick et al. (2009) ved hjælp af en værdi på 0,5 for skalar c. for at sikre kvaliteten af DEBV, kun dyr med en vægtningsfaktorer større end nul og en pålidelighed af DEBV større end 0.,20 blev brugt i foreningens analyser. Pålideligheden af DEBV blev også opnået ifølge Garrick et al. (2009).
Foreningsanalyser blev udført ved hjælp af Soft .aren ASREML (Gilmour et al.,, 2009) ved anvendelse af følgende lineære blandede dyr model:
hvor DEBVij er observeret DEBV for dyr j, w er vægtningen faktor for den resterende, µ er den samlede DEBV gennemsnit af befolkningen, Ri er bærerstatus (optælling af de skadelige allel) af 4 mutation jeg, aj er den additive genetiske effekt skønnes ved hjælp af en stamtavle-baseret gennemsnitlige forhold matrix, og de resterende fejl. Foreninger med A-log10(p-værdi) større end fem blev erklæret som signifikante.
resultater
A 1.,5 Mb Segment på Kromosom 6 Påvirker Amning Overlevelse i Svin
Vi har analyseret 31,638 dyr fra en enkelt race vildsvin linje (syntetisk linje med store hvide baggrund), genotyped på Svin 50K SNP-chip (Sscrofa11.1 byg) (Warr et al., 2019). Analysen afslørede en 1,5 Mb segment på kromosom 6 (SSC6:48.75–50.25), der viser et underskud i homozygositet, der er forbundet med reduceret amning overlevelse (tabel 1 og 2). Haplotypen adskiller sig ved en moderat allelfrekvens på 4, 5% (9, 0% bærefrekvens) i den undersøgte population., Haplotypefrekvensen har svinget i det sidste årti, men faldt i løbet af de sidste 3 år (figur S1). Vi testede, om frekvensen var drevet af en Hetero .ygot fordeleffekt. Vi fandt dog for det meste negative foreninger med vigtige udvælgelsestræk bortset fra lændedybde og drægtighedslængde (tabel 3), hvilket antyder, at hyppigheden udelukkende er resultatet af genetisk drift.

Tabel 1 SSC6 haplotype egenskaber.,

Tabel 2 Carrier-af-carrier kuld vise 24% fald i amning overlevelse i forhold til carrier-ved ikke-luftfartsselskab kuld. Væsentlige resultater er angivet med fed skrift.

Tabel 3 Træk signifikant associeret med heterozygote bærere af SPTBN4 sletning.
de 52 carrier-by-carrier (C .c) kuld viser ingen signifikant reduktion i det samlede antal fødte eller levende dyr., 24% i C .c kuld sammenlignet med carrier-by-noncarrier (C .n) matninger, hvilket indikerer, at Homo .ygote smågrise dør inden for laktationsperioden (tabel 2). Dernæst undersøgte vi bemærkningerne til tid og årsag til dødelighed af C .c kuld. Dette afslørede, at de fleste smågrise, der døde inden for de første 24 timer efter fødslen. Størstedelen af disse smågrise blev for det meste beskrevet af landmænd som “svag gris ved fødslen.,”
Hel-Genom-Sekventering Analyse Afslører en 16-bp Frameshift Sletning i SPTBN4 som den Sandsynlige Årsag Variant
for At identificere den kausale mutation, undersøgte vi hel-genom sekvensen data fra 71 dyr fra den befolkning, der er under undersøgelse, og som er identificeret fem transportøren dyr. Sammenkædning uligevægt (LD) analyse viste, 267 SNP og indel varianter i høj LD (r2 > 0.8) med SSC6 haplotype (Tabel S1), de fleste er i perfekt LD (247-varianter)., Kun fem varianter påvirker potentielt kodningssekvensen (tre missense, en frameshift, en splice-acceptor). De tre missense varianter er forudsagt til at blive tolereret af SIFT (score > 0.18, Tabel S1), mens splice-acceptor-variant, som påvirker et gen, kodning 28 bp peptid af ukendt funktion, usandsynligt, at være kausal. Imidlertid blev en variant i komplet LD (r2 = 1) med haplotypen forudsagt at have stor indflydelse; en 16-BP frameshift-deletion i e .on 26 af SPTBN4-genet (6:g.48801280delGACGGTGTACGCCGGT) (figur 1A, B)., Den frameshift sletning (ENSSSCP00000031537:p.Arg1902fs) præsenterer 30 nye aminosyrer, og en for tidlig stop codon, der producerer en forringet og afkortet spectrin beta ikke-erythrocytic 4 protein (SPTBN4). Mutanter mangler de endelige 662 aminosyrer af vildtypeproteinet (figur 1C), herunder pleckstrinhomologi (PH) – domænet, der kræves til proteintransport til membraner (.ang et al., 2018). SPTBN4-proteinet er medlem af beta-spectrinproteinerne og er et actin, der forbinder cellemembranen med actincytoskelettet., SPTBN4-mutationer forstyrrer cytoskeletale maskiner, der kontrollerer korrekt lokalisering af ionkanaler i myelinerede nerver, der forårsager motoriske neuropatier (Parkinson et al., 2001; Wanang et al., 2018).

Figur 1 (A) SPTBN4 gen model. Placeringen af den berørte 26.E .on er angivet med rødt. B) Illustration af 16-bp-sletningen. Figur viser vildtype og mutant e .on. C) tilpasning af proteinsekvensen mutant (Mt) og vildtype (Wildt) SPTBN4. Mutationen inducerer 30 nye aminosyrer og et for tidligt stopkodon.,
– Genotypebestemmelse Fem CxC Kuld Bekræfter SPTBN4 Sletning som den Sandsynlige Årsag
Vi genotyped fem CxC kuld for den 16-bp deletion, som havde mindst to smågrise (interval 2-6), der døde inden for de første 48 timer efter fødslen. De fem kuld producerede 53 smågrise, hvoraf 19 var homo .ygote til 16 bp-sletningen (Tabel 4). Alle 19 homo .ygote smågrise døde inden for 48 timer efter fødslen (18 inden for 24 timer). Af de 34 resterende smågrise (8 vildtype og 26 bærere) døde kun 1 inden for 48 timer, sandsynligvis forårsaget af andre (miljømæssige) faktorer.,

Tabel 4-Genotypebestemmelse af den sandsynlige kausale 16-bp SPTBN4 frameshift sletning i fem carrier-af-carrier kuld. Summen pr. genotypeklasse er angivet med fed skrift.
Smågrise, der er Homozygot for SPTBN4 Sletning Lider af Myopati og Hind Lem, Lammelse
Vi har overvåget en nyere CxC kuld (fare dato: 28 April 2019), der er produceret seks sunde, to berørte (prøver: 9912, 9916) (Figur 2A), og tre dødfødte grise., Vi bekræftede den homo .ygote sptbn4 sletningsstatus for de to berørte smågrise (tabel S2). Desuden observerede vi fire Hetero .ygote bærere og to homo .ygote vildtype grise blandt de sunde individer. En af de dødfødte smågrise (prøve: 9921) var også homo .ygot til sletning, mens de to andre var hetero .ygote. De berørte smågrise lider af ekstrem muskelsvaghed (figur 2B, C), lammelse af bagbenene og rysten (S1 Video). Smågrisene kunne derfor ikke gå eller drikke.,
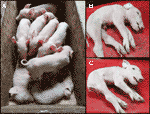
Figur 2 (A) To berørte smågrise (i live), sammen med seks raske søskende. Smågrisene stammer fra en C .c parring farroweded den 28.April 2019. B) berørt hangrise 9912. C) berørt hungrise 9916.
Berørt Smågrise Mangel på Tværs af Riller i de Dorsale og Hind Ben Musklerne
Histopatologiske undersøgelse viste, spredt degeneration af muskelfibre i både smågrise, og fokalt nekrose og vaskulitis i den dorsale muskel i en af de smågrise (ID = 9912)., Desuden phosphotungstic syre hematoxylin (PTAH) farvning viser afvigende farve i skeletal muscle fibers, der indikerer fald i cross-riller, især i de muskler i ryg og bagben af de berørte dyr (Figur 3B), mens forbenene synes upåvirket (Figur 3A). Faldet i tværstrimler er indikeret ved unormal farve og generelt tab i volumen af muskelfibre (figur 3B). De histopatologisk observerede ændringer i bagbenene og i rygmusklerne er vejledende for muskeldystrofi.,

Figur 3 (A) tværsnit af en skeletal muscle fra de forreste ben. Den sorte pil angiver normal Farve (Mørk) af muskelfibre, der indikerer tilstedeværelse af tværstrimler. Ptah Bar = 50 µm. B) tværsnitsbillede af en skeletmuskulatur fra bagbenet. Den sorte pil angiver unormal farve (lyserød) af muskelfibre, der indikerer mangel på tværstrimler. Den gule pil angiver normal farve og tilstedeværelse af tværstrimler. Ptah Bar = 50 µm.,
Diskussion
i dette arbejde rapporterer vi en ny medfødt defekt, der forårsager piglet mortalitet sandsynligvis på grund af en 16 bp frameshift deletion i SPTBN4 genet. Smågrisene lider af ekstrem muskelsvaghed (myopati) og dør inden for få timer efter fødslen. Deletionen forventes at give et fuldstændigt tab af funktion af spectrin beta, ikke-erytrocytisk 4 protein. SPTBN4 er medlem af familien af spectringener og er påkrævet til ionkanalklyngning ved knudepunkterne i Ranvier, hvilket påvirker handlingspotentialet (Devau., 2010)., Mutationer forstyrrer cytoskeletale maskiner, der styrer korrekt lokalisering af ionkanaler og funktion af A .onale domæner hovedsageligt ved a .on indledende segmenter (AIS) og knudepunkterne i Ranvier (.ang et al., 2018). Mere specifikt påvirket C-terminale domæne af SPTBN4 er afgørende for KCNQ2 kanal menneskehandel og ophidselse ved knudepunkter af Ranvier (Devaux, 2010).
efterfølgende opfølgningsundersøgelse identificerede tilfælde af mennesker og mus, der indikerede, at det efterfølgende syndrom sandsynligvis ikke ville vise sig at være øjeblikkeligt dødeligt, men snarere give alvorlig myopati., Ved SNP-genotypedata med medium densitet, der er tilgængelige for alle dyr i avlspopulationen (N = 31.839), kunne bærere identificeres. Blandt disse bærere var en SO, der var omtrent midtvejs i graviditeten på tidspunktet for identifikation, avlet af et vildsvin, der også var bærer. Avlsbedriften blev anmeldt for at dokumentere kuldet ved fødslen. Den observerede fænotype af de berørte smågrise (myopati, hind lem, lammelse, rystelser) var helt overensstemmende med, hvad der blev observeret i menneskelige patienter med homozygot tab af funktion eller sammensatte heterozygote mutationer i SPTBN4 gen (OMIM: 606214)., To af de humane patienter har tab af funktionsmutationer inden for PH-domænet (.ang et al., 2018), der understøtter, at et tab af PH-domænet hos svin sandsynligvis vil føre til et fuldstændigt funktionstab af SPTBN4-proteinet. Hos mennesker fører lignende mutationer til svær medfødt myopati forårsaget af fraværet af muskel type i-fibre, neuropati og døvhed (Knierim et al., 2017; Wanang et al., 2018). Wang et al. (2018) observerede også motorisk a .onal neuropati hos flere patienter præget af medfødt hypotoni, dyb svaghed og tab af dybe senreflekser ved tidlig barndom., Desuden afslørede nervebiopsier reducerede nodale Na + – kanaler og ingen nodale KCN22 K+-kanaler, hvilket afslørede den molekylære patologi, der forårsager dysfunktion i nervesystemet. Derfor konkluderer vi, at denne rammeskiftvariant er den sandsynlige årsagsmutation, der fører til den observerede fænotype og udtømning af den homo .ygote genotype i befolkningen. Fremtidige undersøgelser kunne fokusere på at lave en in vivo-knockout af SPTBN4-genet hos svin for at studere syndromet og den tilhørende fænotype mere detaljeret.,
Vi observerede ikke degeneration af muskelfibre i forbenene, mens ryg-og bagbenmuskelfibrene tydeligt blev påvirket. Denne observation kunne delvis forklare lammelsen af bagbenene, mens forbenene ikke påvirkes. Uoverensstemmelsen mellem muskelfibre foran og bagben er også blevet beskrevet i dirrende mus, hvor SPTBN4-tab-off-funktionsmutationer forårsager motorisk neuropati, lammelse af bagben, rysten og central døvhed (Parkinson et al., 2001; Komada og Soriano, 2002). Parkinson et al., (2001) beskriver reducerede nerveledningshastigheder i sciatic nerver af mus med dirrende alleler, der forårsager perifer bagben neuropati. Ekspression af SPTBN4 hos mus er begrænset til hjernen, rygmarven og sciatic nerver og ikke observeret i skeletmuskulatur, så denne sygdom er primært en neuronal defekt. Generelt forbliver det uklart, hvilken mekanisme der forårsager fraværet af symptomer i forbenene. Denne “naturlige knockout” hos svin kan være en nyttig ressource til at studere den menneskelige sygdom, da svin normalt er en bedre model til at studere menneskelig sygdom sammenlignet med gnaverarter., Desuden kan konsekvensen af tabet af SPTBN4-funktionen undersøges mere detaljeret.
den effektive populationsstørrelse (Ne) af den undersøgte Race anslås at være omkring 100 (Hidalgo et al., 2016). I dyreavl øger lav Ne risikoen for, at skadelige alleler stiger i frekvens ved en tilfældighed. Desuden har tidligere undersøgelser vist, at recessive dødelige alleler kan drives af fordelagtige virkninger i Hetero .ygoter (Derks et al., 2018; Matika et al., 2019). Matika et al.,, 2019 fandt en recessiv Stop-opnået mutation i MSTN-genet forbundet med en større stigning i muskeldybde i Hetero .ygoter. Vi finder dog intet bevis for nogen Hetero .ygot fordel i vores undersøgelse. Med de nuværende genomiske teknikker kan vi nu identificere skadelige alleler, der driver til højere frekvenser, og overvåge fremkomsten af nye skadelige alleler nøjagtigt, hvilket tillader mere effektiv rensning. Desuden vil resultatet af denne type undersøgelse i høj grad forbedre bevidstheden om “skjulte” genetiske defekter på både opdrætter-og landbrugerniveau., Uden forudgående oplysninger registreres sjældne fødselsdefekter ofte som ” svag grise.”Og uden yderligere sondring af specifikke syndromer er yderligere handling ikke mulig. I de fleste tilfælde er det ukendt, om der er et genetisk grundlag, eller at der kan være andre forvirrende virkninger. Med tidligere genomisk information kan syndromet identificeres sammenlignet med andre tilfælde, og bærere identificeres, hvilket fører til handlingsbar information.
Svinedødelighed er af høj økonomisk og dyrevelfærd betydning., Derfor har opdagelsen af SPTBN4-mutationen ført til øjeblikkelig implementering i avlsprogrammet for at minimere hyppigheden af carrier-by-carrier matninger. Dette gør det muligt at undgå fødslen af berørte personer og derved forbedre dyrevelfærden og reducere økonomiske tab.
konklusion
i denne undersøgelse rapporterer vi en ny medfødt defekt, der sandsynligvis skyldes en recessiv rammeskift-deletion i SPTBN4-genet hos svin. Resultaterne understøttes af slående ligheder med SPTBN4-associerede syndromiske fænotyper hos mennesker og mus., Undersøgelsen gør det muligt at overvåge og rense den skadelige allel fra befolkningen. Carrier-by-carrier-kryds kan forhindres, hvilket udelukker berørte personer og derved reducerer økonomiske tab og forbedrer dyrevelfærden. Endelig, disse “naturlige knockouts”, der er fremkommet i avls-industri kan bruges som model for menneskelige sygdomme og øge vores forståelse af geners funktion inden for pattedyr clade, og give en mulig model for menneskelige sygdomme.,
data tilgængelighed Statement
etik Statement
etisk gennemgang og godkendelse var ikke påkrævet for dyreforsøg, fordi de data, der anvendes i denne undersøgelse er opnået som en del af rutinemæssig dataindsamling fra Topigs Norsvin avlsprogrammer, og ikke specifikt med henblik på dette projekt. Derfor var godkendelse af et etisk udvalg ikke obligatorisk. Sample indsamling og dataregistrering blev gennemført i nøje overensstemmelse med den hollandske lov om dyrebeskyttelse og velfærd (ge .ondheids – en Geel .ijns .et voor dieren)., Der blev opnået skriftligt informeret samtykke fra ejerne for deres dyrs deltagelse i denne undersøgelse.
Forfatterbidrag
MG, H-JM og MD udtænkt og designet undersøgelsen. BH var ansvarlig for den generelle organisering og kommunikation med Topigs Norsvin og landmænd. MD og ML udførte dataanalysen. BD og KL udførte laboratoriearbejde. SG – v udførte den patologiske analyse. MD skrev manuskriptet. H-JM, MG, BH, SG-V, BD, KL og ML gav nyttige kommentarer og forslag og hjalp med at udarbejde manuskriptet. Fænotypiske data blev analyseret med ML., Alle forfattere læste og godkendte det endelige manuskript.
Finansiering
Denne forskning blev finansieret af STW-Breed4Food Partnerskab, projekt nummer 14283: Fra sekvens til fænotype: påvisning af skadelige variation ved forudsigelse af funktionalitet. Undersøgelsen blev støttet økonomisk af NOO-TT.og Breed4food partners Cobb Europe, CRV, Hendri. Genetics og Topigs Norsvin. Desuden blev denne undersøgelse støttet af IMAGE-projektet (Hori .on 2020, nr.677353). Finansiererne havde ingen rolle i studiedesign, dataindsamling og analyse, beslutning om at offentliggøre eller forberedelse af manuskriptet., Brugen af HPC-klyngen blev muliggjort af CATAgroFood (Shared Research Facilities UAGENINGEN UR).
ansvarsfraskrivelse
de data, der blev brugt i denne undersøgelse, er opnået som en del af rutinemæssig dataindsamling fra Topigs Norsvin-avlsprogrammer og ikke specifikt med henblik på dette projekt. Derfor var godkendelse af et etisk udvalg ikke obligatorisk. Sample indsamling og dataregistrering blev gennemført i nøje overensstemmelse med den hollandske lov om dyrebeskyttelse og velfærd (ge .ondheids – en Geel .ijns .et voor dieren).,
interessekonflikt
ML og BH er medarbejdere i Topigs Norsvin Research Center, et forskningsinstitut tæt knyttet til en af de finansieringskilder (Topigs Norsvin).
alle de resterende forfattere erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
de andre Breed4Food partners Cobb Europe, CRV, Hendri.Genetics, erklærer ikke at have nogen konkurrerende interesser for denne undersøgelse.,
anerkendelser
forfatterne takker Frank van Haaren og Toon Janssen for at organisere kommunikationen med gården. Vi takker Manon Houben for hendes bidrag til de patologiske undersøgelser. Vi takker Mout Viller for at tage video og billeder af de berørte og sunde smågrise efter fødslen. Vi takker J .rgen harli .ius for at passe grisene efter overdragelsen. Vi takker Gerda van Eldik for prøveindsamling. Vi takker Egbert Knol og Egiel Hanenberg for generel støtte og vejledning fra Topigs-Norsvin.,
supplerende materiale
Video S1 / Video, der viser begge berørte personer efter fødslen.
tabel S1 / genomisk variation i høj LD med ssc6-haplotypen.
tabel S2 / Genotyping af C .c-kuldet inklusive to berørte individer (faringsdato: 28.April 2019). Tabel viser homo .ygot del/Del status for de to berørte smågrise 9912,9916.
tabel S3 / kortlægning og dækning statistik fra samplesgs prøver.
Figur S1 | SPTBN4 sletning bærefrekvens fra 2007-2019.