Indledning
i Modsætning til dyr, der kan bevæge sig, for at undgå de skadelige omgivelser, den siddende planter udviser en højt udviklet tilpasning til den komplicerede miljømæssige forhold. For at opnå denne dybe tilpasningsevne er kommunikation mellem celler nødvendig. Celle til celle kommunikation i planter involverer robust intracellulær signalbehandling og indviklede intercellulære signalnetværk., Indtil nu er der mindst ni signalering stoffer, som er opkaldt plante hormoner, herunder auxin, brassinosteroid (BR), cytokinin, gibberellins (GA), ethylen, jasmonic syre (JA), strigolactone (SL), abscisinsyre (ABA), og salicylsyre (SA) opdaget (Druege et al., 2016; Verma et al., 2016). De genetiske og fysiologiske undersøgelser har afsløret de kritiske roller og funktionelle mekanismer for disse ovennævnte hormoner i plantevækst og udvikling (Gray, 2004)., Baseret på tidligere undersøgelser, auxin, BR, GA, SL, og cytokinin primært funktion under normale planters vækst og udvikling, mens ABA, ethylen, JA, og SA spiller en vigtig rolle i planternes vækst svar på forskellige biotiske og abiotiske stressfaktorer (Pieterse et al., 2009; Santner et al., 2009; denance et al., 2013). Og også nogle af disse hormoner har to roller, for eksempel spiller ABA også vigtige roller i frøudvikling og dvaletilstand (Seo og Koshiba, 2002)., Selvom hvert hormon spiller dominerende roller i visse aspekter, har mange hormoner overlappede aktiviteter, og interaktionerne mellem forskellige hormoner kontrollerer mange udviklingsaspekter og vækst som reaktion på endogene udviklingsmæssige og eksogene signaler.AU Auxin og BR er to store klasser af vækstfremmende hormoner., BR, en gruppe af plante-specifikke steroid hormoner, som kunne interagere med andre plantehormoner som auxin, cytokinin, ethylen, GA, JA, og SA og regulere en bred vifte af plante vækst og udviklingsmæssige processer, herunder frø, spiring, celle forlængelse, vaskulær differentiering, stomata dannelse og bevægelse, blomstring og mandlig fertilitet (Saini et al., 2015). Interessant nok styres hver af disse processer også af Au .in, hvilket antyder, at disse to hormoner spiller sammen for at kontrollere planteudviklingen., I denne gennemgang vil vi redegøre for det signal transduktion af auxin og BR baseret på de seneste fremskridt og gennemgå den krydstale mellem auxin og BR medieret planters vækst og udvikling.
AU Auxin signalvej
AU Auxin blev først anerkendt som plantehormon på grund af dets rolle i plantetropisme til tyngdekraft eller lysstimuli., Senere auxin var kemisk identificeret som indol-3-eddikesyre og har vist sig at spille væsentlige roller i overflod af anlægget udviklingsmæssige og fysiologiske processer, herunder embryogenese, organogenesis, vaskulær differentiering, rod og skyde udvikling, tropic vækst, og frugt udvikling (Estelle, 2011).
Ved anvendelse af genetisk analyse i Arabidopsis er den molekylære mekanisme, der ligger til grund for AU .in-signaltransduktionen, blevet undersøgt godt. TRANSPORTINHIBITOR RESPONSE1 (TIR1) var den første identificerede nukleare receptor af Au .in (Ruegger et al., 1998; Dharmasiri et al., 2005)., TIR1 koder for et nukleart protein, der tilhører f-BO. – proteinet som en underenhed af SCF E3 UBI .uitin ligase proteinkompleks (Gray et al., 1999, 2002; Hellmann et al., 2003; Quuint et al., 2005), I tillæg til TIR1, er der yderligere tre F-boks proteiner nemlig AUXIN SIGNALERING F MAX Proteiner (AFBs), der viser, auxin-bindende aktivitet og mægle auxin signalering i Arabidopsis (Badescu og Napier, 2006). TIR1 receptor kan interagere med en gruppe af AUX/IAA (auxin/indol-3-eddikesyre), proteiner (Dharmasiri et al., 2003)., AU./IAA-proteiner er negative regulatorer af Au .in-signalering, og der er 29 medlemmer af AU. / IAA kodet i Arabidopsis-genomet. AU. / IAA-proteiner kunne interagere med klassen af transkriptionelle regulatorer, AU .in-responsfaktorer (ARF), for at formidle transkriptionelle reaktioner på Au .in. Under høj auxin niveau, AUX/IAA proteiner, der interagerer med TIR1 som coreceptor af auxin, og kan være ubiquitinated af SCFtir1 komplekse og dermed blive forringet gennem ubiquitin–proteasome vej (Gray et al., 2001; Lan Lana et al., 2012)., Efter ødelæggelsen af AUX/IAA repressors, den auxin transkriptionel tilsynsmyndigheder ARFs, som omfatter 23 medlemskaber kan blive frigivet fra AUX/IAA undertrykkelse og dermed mægle auxin svar ved aktivering eller undertrykkelse af target gener (Guilfoyle og Hagen, 2007). De forskellige sæt f-BO. – protein og Au./IAA eller ARFs udleder kompleksiteten under au .in-signaltransduktion (Goh et al., 2012; Guilfoyle, 2015; Salehin et al., 2015).,
den koordinerede virkning af Au. / IAA-transkriptionelle undertrykkere og ARF-transkriptionsfaktorer producerer komplekse genregulerende netværk, som også blev rapporteret i Physcomitrella (Lavy et al., 2016). For nylig, blev det konstateret, at CULLIN1 (CUL1), ændring af SCF interagerer med TIR1 og dermed regulerer TIR1 substrater stabilitet og auxin-signalering (Wang et al., 2016). Interaktionen mellem TIR1 og Au./IAA påvirkes også af den rumlige konformation af Au./IAAs, kontrolleret af en cyclophilinisomerase LRT2 i ris (Jing et al., 2015)., HEAT SHOCK FAKTOR 90 (HSP90) og co-chaperone SGT1, henholdsvis, interagerer med TIR1 og dermed regulerer TIR1 stabilitet, som påvirker samspillet mellem TIR1 og Aux/IAA og auxin-signalering (Wang et al., 2016).
Udover TIR1-afhængige kanoniske auxin-signalering vej, auxin er for nylig blevet rapporteret, at fremkalde en bred vifte af udviklende reaktioner gennem en ikke-kanoniske auxin-signalering mekanisme., I denne ikke-kanoniske auxin sensing proces, ARF3/ETTIN styrer geners udtryk gennem interaktion med proces-specifikke transkriptionsfaktorer, som meget beriger auxin-medieret plante udviklingsmæssige mangfoldighed (Simonini et al., 2016, 2017).
br-signalvejen
BRASSINOSTEROID blev først opdaget i pollen på grund af dets evne til at fremme celleforlængelse. Senere blev det konstateret, at br spiller roller i en lang række plantevækstaspekter og kan reagere på biotiske og abiotiske belastninger., I dag blev br-signaltransduktionsvejen stort set afklaret ved kombinationer af forskellige metoder, herunder molekylær genetik, biokemi, proteomik og genomik osv. Det celle-overflade-kinase BRASSINOSTEROID INSENSITIVE1 (BRI1) blev identificeret som den receptor af BR, som kan binde sig til den ekstracellulære domæne af BRI1 og aktivere sin kinase aktivitet og dermed skifte til en signal-kaskade at regulere transkription (Li og Chory, 1997; Wang et al., 2001; Kinoshita et al., 2005; Kim and andang, 2010; Clouse, 2011; Hothorn et al., 2011; hun et al., 2011; Oh et al., 2012)., Efter opfattelse af BR interagerer BRI1 med co-receptor BRI1-associeret KINASE 1 (BAK1) og dets homolog somatiske EMBRYOGENESISRECEPTORKINASER (SERKs) for at danne et mere aktivt BR-receptorkompleks (Li et al., 2002; Nam og Li, 2002; Wanang et al., 2005; Tang et al., 2008; Gou et al., 2012). Aktiveret BRI1 phosphorylates to substrater af plasma-membran-forankret receptor-lignende cytoplasmatisk kinaser: BRASSINOSTEROID-SIGNALERING KINASES1 (BSK1) og KONSTITUERENDE DIFFERENTIERET VÆKST1 (CDG1) (Tang et al., 2008; Kim et al.,, 2011), hvilket igen phosphorylates en PP1-type fosfatase opkaldt BRI1-SUPPRESSOR1 (BSU1) for at aktivere BSU1, der fører til BSU1 dephosphorylation og inaktivering den GSK3-som kinase BRASSINOSTEROID INSENSITIVE2 (BIN2). Kinaseaktiviteten af BIN2 hæmmes også af HISTONDEACETYLASE HDA6, som interagerer og deacetylerer ved K189 af BIN2., Når BR niveauet er lavt, BRI1 er inaktiv på grund af sin negative regulator, BRI1 KINASE INHIBITOR-1 (BKI1) og protein phosphatase 2A (PP2A), mens BIN2 phosphorylate to BR homologe transkriptionsfaktorer, BRASSINAZOLE RESISTANT1 (BZR1) og BZR2 (også kaldet BES1 for BRI1-EMS-SUPPRESSOR-1) (Han et al., 2002; Wanang et al., 2002; Yin et al., 2002; Mora-Garcia et al., 2004; Kim et al., 2009, 2011; Kim og Kimang, 2010). Når br-niveauerne er høje, inaktiveres BIN2, og B .r1 og B .r2 dephosforyleres af PP2A og bevæger sig ind i kernen for at ændre ekspressionen af tusinder af BR-Responsgener (He et al.,, 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011).
synergien mellem BR-og Au .in-signalering
AU .in-og BR-signalveje spiller forskellige roller, men de viste også synergistiske og indbyrdes afhængige interaktioner i en lang række udviklingsprocesser. For eksempel kan både AU .in-og BR-signaler fremme celleudvidelse og kan interagere synergistisk for at fremme hypokotylforlængelse (Nemhauser et al., 2004)., Svaret fra en af de to veje til fremme af hypocotylforlængelse kræver den anden funktion og indbyrdes afhængighed mellem BR-og Au .in-veje (Nemhauser et al., 2004). Auxin øget hypocotyl længde i wild-type planter, men ikke i BR-ufølsom over mutant bri1-116, og dette auxin-ufølsom fænotype af bri1-116 blev undertrykt af den dominerende gain-of-function mutant bzr1-1D, hvilket indikerer, BR eller aktiv BZR1 er nødvendig for auxin fremme af hypocotyl forlængelse., Det er blevet konstateret, at BR signalering konvergerer med SUPPRESSOR AF PHYTOCHROME B4-3 (SOB3) til at styre celle forlængelse og hypocotyl vækst gennem regulering af auxin induceret LILLE AUXIN OP RNA19 (SAUR19) udtryk (Favero et al., 2017). På den anden side, auxin reguleret transskription faktor LILLE ORGEL STØRRELSE 1 (SMOS1) er for nylig blevet fundet til at styre celle ekspansion gennem den direkte interaktion med SMOS2/DLT, et medlem af den GRAS familie af transkriptionel co-myndigheder, der spiller en positiv rolle i BR signalering i ris (Kim et al., 2009; Tong et al., 2012; Hirano et al., 2017)., Auxin relaterede mutanter som iaa3 og arf6/arf8 var mindre følsomme over for BR, end det var wild-type for hypocotyl forlængelse, og afskaffet overfølsomhed over for bzr1-1D til auxin, hvilket tyder på, BR og BZR1 fremme af hypocotyl forlængelse kræver ARF6/8. Den genom-brede ChIP-se.-analyse afslørede, at ARF6 deler et stort antal genomiske mål (omkring 50%) med B .r1 og den lys/temperaturregulerede transkriptionsfaktor PIF4 ved CHIP-se. analyser (Oh et al., 2014)., B .r1 og PIF4 interagerer med ARF6 og aktiverer delte målgener ved at binde til delte målgener kooperativt under hypocotyls forlængelse (Oh et al., 2014) og mange af disse overlappende målgener koder for cellevægsproteiner involveret i celleudvidelse.
Brassinosteroid og Au .in spiller også vigtige roller i vedligeholdelsen af root apical meristem (RAM) (Durbak et al., 2012). Rammen består af en lille gruppe sjældent opdelte celler kendt som quiuiescent center (.c), omgivet af stamceller, der giver anledning til de forskellige tootvævstyper., Vedligeholdelsen af rodstamcellepopulationen reguleres af HOMEUSCHEL-relateret HOMEOBO.5 (.o .5) (Sarkar et al., 2007). Wo .5 er begrænset til QC ved au .in-signalering og letter korrekt ekspression af PLT-generne (Aida et al., 2004; Ding og Friml, 2010). Mutationer i BR-receptor-genet BRASSINOSTEROID UFØLSOM 1 (BRI1) resultere i abnorme celle cyklus progression i RAM og forårsage en mindre RAMs (Gonzalez-Garcia et al., 2011; Hacham et al., 2011). AU .in er kendt for at stimulere biosyntesen af br (Chung et al.,, 2011), men aktiviteten af BR påvirker ikke ekspressionen af PIN-gener (Hacham et al., 2011). Rodspidsfænotyperne af BR-mutanter viser ikke det samme som au .in-mutanterne (Gon .ale. – Garcia et al., 2011), hvilket indikerer, at BR handler på RAM uafhængigt af Au .in.
Brassinosteroid-og Au .insignaler kræves også synergistisk i den radiale mønsterdannelse af vaskulære bundter (ibanes et al., 2009)., Ved kombinationer af matematisk modellering og biologiske eksperimenter, auxin maxima, der er etableret af asymmetrisk auxin polar transport, men ikke ændringer på auxin niveauer er vigtige for positionering af vaskulære bundter. BR-signalet blev vist at tjene som et fremmende signal for antallet af celler i den provaskulære ring, som er i overensstemmelse med AU .in ma .ima. Således er etableringen af periodisk arrangement af vaskulære bundter i skuddet under den koordinerede virkning af disse to plantehormoner (ibanes et al., 2009)., Begge signaler er også involveret i plantens rodudvikling, og interaktionen mellem BR og Au .in formidles af BREVIS radi. (br.) under denne proces. BR.er vigtig for den hastighedsbegrænsende biosyntese af BR og BR eksogen applikation kan redde br.-mutante defekter. Desuden, auxin-responsive genekspression er globalt nedsat i brx mutant, og udtryk for BRX er stærkt foranlediget af auxin og undertrykt af BR, hvilket indebærer, at BR biosyntese og auxin signalering er tilsluttet via et feedback loop, der involverer BRX under root udvikling (Mouchel et al., 2006).,
Brassinosteroider og Au .in spiller også synergistiske roller under lateral rodudvikling. BRs fungerer hovedsageligt ved initiering af lateral rod primordia, mens AU .in er påkrævet til både initiering og fremkomststadier af lateral roddannelse (Casimiro et al., 2001; Bhalerao et al., 2002; Benkova et al., 2003; Bao et al., 2004). Under disse processer øger BRs LRP-initiering ved at fremme akropetal AU .intransport i roden, men ikke ved at påvirke endogent IAA-niveau (Bao et al., 2004)., Alle disse rapporter tyder på, at krydstale mellem BR og Au .in spiller en vigtig rolle i reguleringen plantevækst og udvikling.
BR Regulerer Auxin Signalering
Udover den gensidige afhængighed og samarbejde af auxin og BR signaler i løbet af planternes udvikling, BR kunne mægle auxin signal vej på flere niveauer. BZR1 interagerer med ARF proteiner direkte mod flere auxin signalering komponenter og gener, der er involveret i auxin stofskiftet, såsom transport og signaler, herunder AUX/IAA, PINs, TIR1, og ARFs, osv. (Sun et al ., 2010)., Det blev konstateret, at Aux/IAA proteiner, der er involveret i BR svar og iaa7/axr2-1 og iaa17/axr3-3 mutanter viste afvigende BR følsomhed og afvigende BR-induceret genekspression i et organ, der er afhængige måde (Nakamura et al., 2006). Eksogen brassinolid (BL)-behandling kunne inducere ekspressionen af Au .in-responsive gener, såsom IAA5, IAA19, IAA17 osv. og udtrykket af ovenstående gener er nedreguleret i BR biosyntesereaktioner mutant de-etiolated2 (det2), hvilket indikerer, at funktionelle BR biosyntesen er dels nødvendig for auxin-afhængige genekspression (Nakamura et al.,, 2003; Kim et al., 2006). Derudover påvirker BR også AU .in Flo.ved at regulere ekspressionen af Au .in eksportører som PIN4 og PIN7 (Nakamura et al., 2004). I løbet af anlægget gravitropism svar, BRs kunne øge polar ophobning af auxin eksportør PIN2-i roden meristem zone, og dermed påvirke den omfordeling af auxin fra roden spids mod forlængelse zoner og resultere i forskellen på IAA niveauer i både øvre og nedre sider af rødder til at fremkalde plante gravitropism., Under denne proces spiller BR aktiveret ROP2 en vigtig rolle i modulering af den funktionelle lokalisering af PIN2 gennem regulering af montering / genmontering af F-actins (Li et al., 2005). Yderligere undersøgelser viste, at nedsat BL-opfattelse og / eller koncentration kunne inducere CYP79B2, genet, der koder for et en .ym, der omdanner tryptophan til indol-3-acetaldo .im og således påvirker distributionen (Kim et al., 2007).
derudover blev det konstateret, at br-signalet kunne regulere AU .in-signaludgang ved sin negative regulator GSK3 kinase BIN2., AU .in-responsfaktoren ARF2 blev identificeret som et BIN2-interagerende protein i en gær-to-hybridskærm, og kinase-assay viste, at BIN2 kunne phosphorylere ARF2. Phosphorylering af ARF2 resulterer i tab af dets DNA-bindingsevne og undertrykkelsesaktivitet af målgenerne (Vert et al., 2008). ARF2 er en B .r1-målgener, og dens ekspression reduceres ved BR-behandling (Sun et al., 2010)., Derudover BIN2 kan phosphorylate ARF7 og ARF19 til at undertrykke deres interaktion med AUX/IAAs og derved øge transkriptionel aktivitet på deres target gener LATERAL ORGEL GRÆNSER-DOMAIN16 (LBD16) og LBD29 til at regulere lateral root organogenesis (Cho et al., 2014). BR spiller imidlertid en mindre rolle under denne proces, og BIN2 er under kontrol af det TRACHEÆRE ELEMENTDIFFERENTIERINGSHÆMMENDE faktor (tdif)–tdif-RECEPTOR (tdr) modul (Cho et al., 2014). Sammen kan BR regulere AU .in reponses ved at påvirke forskellige AU .in signalering komponenter.,
AU .in regulerer br-signalering
på den anden side kan AU .in også regulere br-signalvejen i visse aspekter. Udtrykket af D .arf4, en afgørende hydro .ylase for BR biosyntese til at kontrollere endogent BR-niveau, er Au .inafhængig. AU .inbehandling kunne mærkbart stimulere ekspressionen af D .arf4, og Au .in kunne hæmme bindingen af B .r1 til promotoren af D .arf4. Induktion af D .arf4 af Au .in kræver AU .in signalvejen, men ikke BR signalvejen (Chung et al., 2011; Yoshimitsu et al., 2011)., CPD-katalyserende C-3-o .idation af br blev aktiveret af BR., en formodet transkriptionsfaktor, der virker nedstrøms for AU .in-signalering (Mouchel et al., 2006). Yderligere undersøgelse i ris viser, at eksogene auxin kan forbedre transskription udtryk niveauer af BR-receptor-genet OsBRI1, hvilket tyder på, at auxin øger BR signalering gennem regulering af BR-receptorer (Sakamoto et al., 2013). Endvidere har promotoren af osbri1 et upstream AU .in-response element (Aurere) – motiv, som er målrettet mod ARF-transkriptionsfaktorer., Desuden indikerer mutantundersøgelser, at ved mutation af Au .re afskaffes induktionen af ekspression af OsBRI1 af Au .in, og også ekspressionen af OsBRI1 er nedreguleret i arf-mutant (Sakamoto et al., 2013). Det er rapporteret, at OsARF19 binder til promotoren af OsBRI1 og positivt regulerer dets ekspression, som derefter aktiverer br-signaleringen (.hang et al., 2015). BES1 kan binde sig til fortaler for SMÅ AUXIN-OP RNA 15 (SAUR15) og mægle BR tidlig indsats gen i Arabidopsis, og denne binding kan være forstærket af auxin behandling (Walcher og Nemhauser, 2012)., Samlet set kan AU .in også påvirke br-responser og Br-reguleret plantevækst og-udvikling.
Afsluttende Bemærkninger og Perspektiv for Fremtiden
i Løbet af de sidste næsten fire årtier, undersøgelser af auxin-BR vej interaktioner har tiltrukket flere og flere forskeres interesse. Apparatet af fysiologiske, molekylære, genetiske og biokemiske værktøjer har i høj grad uddybet vores forståelse af dette problem., Baseret på de tidligere undersøgelser er BR og Au .in involveret synergistisk i flere planteudviklingsprocesser, herunder: hypocotylforlængelse, udvikling af vaskulære bundter, rodudvikling og tropismer osv. AU .in og BR ‘ s indbyrdes afhængighed og samarbejde er komplicerede og involverer adskillige processer på molekylært niveau ved at dele de samme målgener og regulere hinanden gensidigt på flere niveauer (Figur 1).
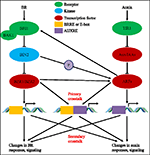
FIGUR 1. Model af Au .in-brassinosteroid (br) krydstale., I Arabidopsis genkendes opfattelsen af BR-og Au .in-signal af henholdsvis BRI1-og TIR1-receptorer. BR binder til det ekstracellulære domæne af BRI1 og fremmer det interagerer med co-receptor BAK1 for at danne et mere aktivt BR-receptorkompleks, hvilket igen fører til dephosphorylering og inaktivering af BIN2. Inaktivering af BIN2 føre til dephosphorylation af to BR homologe transkriptionsfaktorer BZR1 og BZR2, som bevæger sig ind i kernen for at aktivere transskription af gener, som indeholder BRRE eller E-boks i deres promotor region., BIN2 kan også phosphorylere ARF7 og ARF19 for at undertrykke deres interaktion med AU./IAAs og derved forbedre den transkriptionelle aktivitet på deres målgener. TIR1 modtager AU .in signalering og interagerer med AU./IAA proteiner som co-receptor af Au .in. AUX/IAA så er forringet gennem ubiquitin–proteasome vej, og auxin transkriptionel tilsynsmyndigheder auxin svar faktorer (ARFs) er frigivet fra AUX/IAA undertrykkelse og aktivere transskription af gener med auxin responsive elements (AUXRE) i deres lovgivningsmæssige område., Nogle ARF ‘ er kan også binde sig til promotoren af BRI1 og regulerer dets udtryk positivt, som derefter aktiverer br-signaleringen. Primær krydstale sker ved aktivering af gener, der indeholder både BRRE/E-bo.og Au .re i deres promotorregion, hvilket giver begge signalveje mulighed for direkte at regulere transkription. Sekundær krydstale forekommer gennem ekspression af gener, der enten er Au .in eller BR lydhør, men hvis aktiviteter styrer ekspression af gener, der regulerer respons og signalering af andre hormoner.,
Phosphorylering forordning spiller en afgørende rolle i BR signalering vej, især i opfattelsen proces, BR opfattes gennem BRI1 kinase receptoren og BAK1 kinase co-receptorer, og i sidste ende styrer BR reguleret gen udtryk gennem at påvirke nedstrøms transkriptionsfaktorer, som BES1/BZR1 aktiviteter (Han et al., 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011). Imidlertid synes allestedsnærværende regulering afgørende for AU .in-signalering., Når auxin, der binder sig til TIR1 receptor, der virker som en ubiquitin E3-ligase, den aktiverede TIR1 E3-ligase ubiquitinates AUX/IAA proteiner, fører til nedbrydning af disse repressors og de-har fortrængt ARF transkriptionsfaktorer, og i sidste ende forårsager auxin reguleret gen-ekspression mønster ændringer og vækst svar (Gray et al., 1999, 2002; Hellmann et al., 2003; Quuint et al., 2005). Da det har vist sig, at BIN2-kinase, som er kendt funktion i BR signalering, kunne phosphorylate og forbedre aktiviteterne i ARFs som ARF2 og ARF7 (Vert et al., 2008; Cho et al.,, 2014), vil det være interessant at teste, om kinaser som BIN2, der er involveret i BR signalering, kan også interagere med andre auxin signalering komponenter såsom TIR1 receptor-eller AUX – /IAA repressors, og indflydelse TIR1 E3-ligase aktivitet eller AUX/IAA protein stabilities. På den anden side skal allestedsnærværende rolle i BR-signalering også løses, især hvis TIR1 E3-ligase direkte kunne interagere med BR-signaleringskomponenter og regulere deres proteinstabiliteter.,
derudover er det observeret, at Au .in regulerer plantevækst og udvikling på en væv eller cellulær afhængig måde ved hjælp af Au .in-respons DR5 og andre AU .in-journalister. De forskellige transkriptionelle udgange afhængigt af den cellulære og miljømæssige kontekst (Clark et al ., 2014; Etchells et al., 2016; Lavy et al., 2016)., Selvom spatiotemporal BR signalering har vist sig at styre rod vækst gennem antagonistiske indsats med auxin (Chaiwanon og Wang, 2015), det er stadig ukendt, hvis væv eller cellulære BR-signalering, der kan visualiseres ved pBZR1:BZR1-YFP, er også vigtigt at kontrollere andre processer ud over root udvikling. Endvidere er generering af et detaljeret vævs-eller kælderkort over AU .in-og BR-distributioner i øjeblikket muligt ved hjælp af fluorescensaktiveret cellesortering eller lasermikrodissektion i kombination med højopløsningsgenekspressionsanalyse., Dette vil i sidste ende fører til adresse, hvis AU .in crosstalks med BR i et væv eller cellulær måde.
Forfatterbidrag
alle forfattere var involveret i skrivningen af dette gennemgangsmanuskript. For mere information om, hvad der udgør forfatterskab, henvises til vores forfatter retningslinjer.,
Finansiering
Dette arbejde er støttet af tilskud fra National Natural Science Foundation of China (Projekter 31500229 og 31770305), Shandong-Provinsen Natural Science Foundation of Major Basic Research Program (2017C03), Shandong Provinsen Midler for fremragende Unge Forskere (2014JQ201408).
Erklæring om interessekonflikt
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
Clouse, S. D. (2011)., Brassinosteroid signaltransduktion: fra receptorkinaseaktivering til transkriptionelle netværk, der regulerer planteudvikling. Plantecelle 23, 1219-1230. doi: 10.1105/tpc.111.084475
PubMed Abstract | CrossRef Fuld Tekst | Google Scholar
Estelle, M. (2011). Auxin Signalering: Fra Syntese til Systembiologi; Et Emne Samling fra Cold Spring Harbor Perspektiver i Biologi. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Google Scholar
Guilfoyle, T. J. (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar