genet p53 blev først opdaget i 1979. Et protein blev identificeret i simian virus 40-transformerede musceller (SV40) ved immunpræcipitation med anti-t serum; dette protein blev kaldt protein p53 . Samme år fandt Kress og andre forskere en ny klasse af proteiner med en molekylmasse fra 50-60kDa. Denne type protein blev derefter identificeret som p53 . Proteinet p53 kan også identificeres fra forskellige transformerede cellelinjer ved immunpræcipitation., Lane og Lin .er fik også et lignende resultat i 1979. Andre beviser for identifikation af p53 er, at p53 blev udtrykt i alle testede transformerede museceller; disse test inkluderer kemisk inducerede sarkomer, transformerede fibroblaster og leukæmier, mens p53 i normale celler ikke blev udtrykt. Derudover blev der påvist et højt niveau af p53 i de fleste transformerede celler, uanset hvordan cellerne blev transformeret, enten spontant eller ikke-spontant ., Det skyldtes i vid udstrækning den øgede stabilitet af p53, men i F9-embryonale carcinoa-celler udtrykte det et højt niveau af p53, dette skyldtes mængden af oversat p53 mRNA .
efter at proteinet p53 var blevet opdaget i 1979, blev det populært at analysere det. Men på det tidspunkt, da det var et nyopdaget protein, og der ikke var et tidligere navn til det, brugte forskellige institutioner forskellige navne og offentliggjorte papirer med forskellige navne., For at løse dette problem mødtes forskere fra forskellige forskningsgrupper i forskellige lande i 1983 under det 1.Internationale p53-værksted, der blev afholdt i O .ted, Storbritannien, for at diskutere en fælles nomenklatur for dette nyopdagede protein. På dette møde bliver” p53 ” dets navn, og det er blevet brugt siden da. Det blev antaget, at grunden til, at forskere kaldte proteinet p53, er, at molekylmassen af dette protein er 53kDa, der er baseret på dets migration i SDS gel. Senere blev molekylmassen vist sig at være forkert, og den korrekte molekylmasse skulle være 43.,7kDa fordi p53 indeholder en prolinrig region, og denne region kan reducere migrationen af p53 i SDS gel. Men navnet” p53 ” forblev .
i 1980 ‘ erne blev proteinet p53 antaget at være involveret i cellecyklussen såvel som at spille en rolle i DNA-replikation. Senere, i 1982 til 1994, fandt folk, at nogle virale oncoproteiner var i stand til at binde til p53 og danne et kompleks. I 1982, Sarno.et al. fandt, at adenovirus E1b (58kDa) kan interagere med et 54kDa-protein, der er til stede i SV40-transformerede museceller nævnt ovenfor., Ifølge resultaterne af immunologiske specificiteter af T-antistoffer og peptidkortene af 54kDa-proteinet identificeres dette 54kDa-protein som p53 . I samme år, fandt forskerne, at hvis de injicerede det p53 antistof i Schweiziske 3T3 musen celler, at det ville hæmme cellerne ind (S-fasen i cellecyklus, men, under samme situation, p53 antistof ikke påvirke SV40 eller adenovirus induceret DNA-syntese .,
Senere i 1984, har forskere undersøgt effekten af p53 på ikke forvandlet 3T3 fibroblaster; de analyserede syntese sats af protein p53 på forskellige tidspunkter, og fandt, at der i slutningen af G1-fase syntese sats og niveauet af proteinet p53 og dens tilknyttede mRNA stigning. Dette resultat antyder, at proteinet p53 hæmmer celler, der kommer ind i delingsfasen fra interfasen . Malt maltman W et al. gjorde et andet eksperiment i samme år. De behandlede den ikke-transformerede muscelle med UV-lys og UV-mimetisk kemisk kræftfremkaldende 4NOO, og de opdagede et højt niveau af p53., Resultatet viste, at forhøjet ekspression af p53 ikke kun er et symbol, der angiver cellecyklussen, men også vigtigere en komponent, der er involveret i DNA-syntese og celleproliferation . I 1987, når man studerede komplekset af T-antigen af simian-virus 40 og DNA-polymerase a, Gannon og andre forskere fandt en lignende ændring i antigenet, når det var bundet til p53 og polymerase a. De fandt også, at de ved en vis koncentration af de tre komponenter kan danne et specielt trimerisk kompleks, der indbefatter t-antigen, p53 og DNA-polymerase a., Da T-antigen er involveret i viral DNA-replikation og cellulær transformation, indikerer dette resultat, at p53 spiller en rolle i kontrollen af cellecyklussen og DNA-replikation .
som eksperimentet viste ovenfor, har p53 evnen til at udødeliggøre celler. I 1984, Eliyahu d et al. fandt, at p53 og produktet af oncogene myc delte nogle lignende egenskaber. Begge har evnen til at binde til andre proteiner og er involveret i cellecyklussen, og de akkumuleres begge i kerner af transformerede celler ., Bien., Pennica og Oren analyserede aminosyresekvenserne af proteinet p53 og produktet af myc, og de fandt, at de to proteiner viser ligheder i molekylstruktur og placeringen af specielle ladede rester. Derefter foreslog forskere, at p53 kan fungere som en onkogen. Baseret på denne hypotese, Eliyahu d et al. gjorde nogle eksperimenter. Da embryofibroblaster med primær hastighed kan transformeres ved involvering af både myc-produkt og Ha-ras, primære baby rotte-nyreceller kan også transformeres ved samarbejde mellem Ha-ras og adenovirus tidlige region 1A , Eliyahu D et al., besluttet at bruge denne form for biologisk testsystem til at identificere den onkogene funktion af p53. De behandlede normale embryonale celler med p53 og aktiverede Ha-ras. Resultatet viste, at målceller støder på morfologiændringer og producerer høje niveauer af p53, Eliyahu d et al. troede, at omdannelsen af embryonale fibroblaster af p53 og Ha-ras forklarede, at genet p53 er en onkogen ., I 1985 foreslog Jenkins, at p53-genet kan forlænge cellernes levetid, forbedre affektiviteten af transformation ved at omarrangere dens kodende sekvens, der kan forårsage produktion af stabile proteiner .
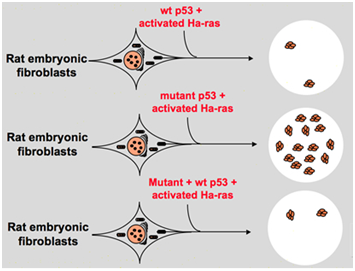
i slutningen af 1980 ‘ erne begyndte forskere imidlertid at indse, at p53 er et tumorundertrykkende gen i stedet for et onkogen. De observerede, at p53 med normal funktion ikke kan påvises i mange af tumorerne og fandt, at det er nødvendigt at miste ekspressionen og funktionen af vildtype p53-gen under celletransformation., Disse øger muligheden for, at vildtype p53-gen kan hæmme neoplastisk progression . Derefter formulerede de en anden hypotese: klongenet p53, der blev brugt i tidligere eksperimenter, indeholder lejlighedsvis dominerende negative mutationer inden for det stærkt konserverede domæne, hvilket fører til modsatte eksperimentresultater . I 1988, ben og andre forskere opdaget en enorm mængde af omarrangeret p53 i murine erythroleukemia cellelinier–dp20-1 og CB3, som er afledt af milten af murine inficeret med ven leukæmi virus ., I 1989 ændrede Eliyahu, der påpegede, at p53 er en onkogen, mening, og han antog, at vildtype p53-gen kan hæmme celletransformation. Eliyahu og andre forskere studerede effekten af vildtype p53-protein kodet af plasmider og mutant p53 på evnen til at fremkalde embryofibroblasttransformation med primær hastighed ved forskellige onkogenkombinationer in vitro. For eksempel mutant p53 plus ras, og myc plus ras., Resultatet viste, at vildtype p53 føre til, at en stor reduktion af den transformerede foci forårsaget af mutant p53 plus ras; mutant p53 viste ingen hæmning på forvandlet foci forårsaget af myc plus ras, mens myc plus ras-medieret transformation er meget følsomme over for udtryk af vildtype p53. Figur 1 viser dette eksperiment kortfattet. Det viste, at sammenlignet med mutant p53 udviser vildtype p53 en åbenlyst hæmmende effekt på celletransformationen. Effekten er positivt relateret til ekspressionsniveauet for vildtype p53 og negativt relateret til ekspressionsniveauet for mutant p53., Dette eksperiment antydede, at vildtype p53 faktisk kan have en modsat funktion sammenlignet med mutant p53 og kan hæmme tumorigenesen . I øjeblikket anerkendes p53 som et tumorundertrykkende gen. Det anslås, at omkring halvdelen af tumorer er forårsaget af p53. Det er et af de hyppigst muterede gener hos mennesker, og det hyppigst analyserede gen rundt om i verden .
i løbet af de første mange år af 1980 ‘ erne var den biokemiske vej for p53 og virkningen af p53-mutation ikke klar. I 1991 fandt Kern og andre forskere, at en 33-basepar DNA-sekvens binder specifikt til vildtype p53 in vitro. De fandt også, at p53-proteinet indeholder to mutationer, der normalt findes i humane tumorer, der ikke kan binde til denne specifikke DNA-region. Så de antog, at funktionen af p53 afhænger af dens evne til at binde specifikke DNA-sekvenser, og denne evne ændres af mutationer, der findes i humane tumorer., De antager også, at denne 33-basepar DNA-sekvens muligvis ikke er den eneste sekvens, der har evnen til at binde specifikt til p53 hos mennesker; det kan dog hjælpe folk med bedre at forstå funktionen af p53 . Senere viste det sig, at p53 spillede en rolle under cellecyklussen, DNA-reparation, differentiering, initiering af apoptose og angiogenese. Rotter V et al. fandt, at p53 up-regulerer differentiering af celler. For eksempel blev et højt niveau af p53-protein påvist i flere nøgletrin under B-celledifferentiering. Forhøjet p53 kan også påvises under spermatogenese., I mellemtiden kan kun et meget lavt niveau af p53-protein påvises i nogle organer hos voksne mus .
i 1990 blev et nyttigt værktøj opdaget lejlighedsvis. Det er en temperaturfølsom mutant af p53, kaldet p53val135. Det kan fungere som en reel vildtype p53 ved temperaturen på 32,5 oC, undertrykke transformation, og det kan også fungere som andre muterede p53 ved temperaturen 37,5 oC eller over 48oC, fremkalde transformation. Derudover styres dens proliferation ved den tilladte temperatur for transformerede celler, der udtrykker p53val135, og denne form for kontrol er reversibel., Ved at bruge denne p53val135-mutant blev vildtype p53 opdaget for at inducere cellecyklusstop ved enten G1 eller G2/M . I 1991, Elisheva et al. fandt, at den temperaturfølsomme p53val135 udførte en anden funktion i den murine myeloide leukæmi cellelinje. Efter reaktivering af p53val135 i et par dage døde alle celler, og denne død udviser nogle egenskaber ved apoptose . Et år senere blev et lignende resultat opnået af Sha.. En vild type p53 blev transficeret til en human colon tumor-afledt cellelinie EB., Cellerne blev undersøgt under lys-og elektronmikroskoper og viste sig at udvise nogle egenskaber ved apoptose . I 1990 blev Scheffner et al. og andre forskere fandt, at E6, der stimulerer ødelæggelse af værtscellen regulerende proteiner, der er kodet af det kræftfremkaldende human papillomavirus type 16 og 18, og det kan danne et kompleks med vildtype p53 in vitro, hvilket igen forårsager nedbrydning af protein p53 .
i 1992 blev et nøgleprotein mdm2 opdaget, fordi det binder tæt sammen med p53, og det hæmmer transaktiveringen medieret af p53., Den molekylære masse af MDM2 er 90kDa, og det danner et kompleks med både muteret og vildtype p53 . I samme år, Livingstone RL et al. undersøgte, om cellen mistede en eller begge kopier af vildtype p53-alleler, og om det var tilstrækkeligt til at forårsage genforstærkning. Genforstærkning blev hovedsageligt påvist i transformerede celler, men ikke i de normale fibroblaster. Resultatet viste, at celler, der mister en kopi af p53-allelerne, fungerer som wildild-type p53, mens celler, der mister begge kopier af wildild-typen p53-alleler, udviser en højere amplifikationsfrekvens . Et andet eksperiment lavet af Yin y et al., viste et lignende resultat .
i 1993 blev et p53-målgen kaldet CDKN1A identificeret. Det koder for proteinet p21, som er en cyclinafhængig kinaseinhibitor, der hæmmer cyclin-CDK2 og CDK1 ved at binde til dem. I 1993, Szekely fandt, at Epstein-Barr virus nuclear antigen 5(EBNA-5) er kodet af Epstein-Barr virus, og det kan inficere humane B lymphoblastoid celle. Et 66 aminosyre langt peptid er ansvarlig for dannelsen af komplekse EBNA-5-p53, punktmutationer af p53 påvirker ikke dets bindingsevne til EBNA-5., Det hæmmer imidlertid dets formationer af komplekser med andre molekyler . I 1994 beskrev Cho og hans medarbejdere først krystalstrukturen af kompleks p53-DNA. Dette DNA-bindende domæne blev også kaldt kernedomænet. Den indeholder rester 102-292, og består af en beta sand .ich. De demonstrerede også den detaljerede struktur af kernedomænet . Også i 1994, Wanang.. et al. interaktionen mellem hepatitis B-virus protein-protein (HB.) og vildtype p53-protein hos mennesker., De fandt, at HB .kan hæmme p53 ‘ s evne til at binde til andet sekvensspecifikt DNA, efter at det er bundet til p53, og det kan også hæmme associeringen af p53 med transkriptionsfaktorer.
i 1997, Honda R et al. første hypotese, at MDM2 kan udløse p53 ubiquitylation og føre til nedbrydning af p53 af en ubiquitin-proteasome system. De påpegede, at MDM2 binder til N-terminal domæne (NTD) af p53 og fungerer som UBI .uitin ligase E3 . Også i 1997 blev to nye familier af proteiner, p63 og P73 opdaget, der deler betydelig homologi med p53., p73, også kaldet tumorprotein 73, kodes af et gen placeret i 1p36. Placeringen slettes ofte i neuroblastom og andre tumorer. p73 kan aktivere p53-målgener og interagerer med p53 . Yang et al. fundet, at genet p63 er placeret i 32727-29, Og det kan påvises i forskellige mus og humane celler. Ligesom p73 kan p63 transaktivere p53-målgener markant, det kan også inducere apoptose. Et kendetegn ved p63 er, at størstedelen af P63 mangler en N-terminus ., I samme år fandt Serrano og kolleger, at primære murinfibroblaster kan transformeres af ras i fravær af p53 eller P16, og inaktive p53 eller P16 kan lette udødelighedsprocessen for humane celler. Disse fund antyder, at p53 spiller en rolle i cellulær senescens . Derefter blev det i 1997 fundet, at p53 spillede en rolle i indledningen af apoptose. Når celler kommer ind i proliferationsfasen, ville telomererne i slutningen af hvert kromosom forkortes efter hver runde af DNA-replikation på grund af ufuldstændig replikation af DNA med enkelt streng i slutningen af DNA-stativet ., Aktiveret tumorundertrykkende gen p53 begrænser antallet af gange celledeling kan forekomme. Wynford TD fandt, at med tabet af funktionen af vildtype p53 undslipper alle fibroblaster fra apoptose. Transaktionsfunktionen af p53 kan også tændes ved apoptose . Wynford TD foreslog, at der er tre muligheder for, hvordan p53 aktiveres. Den første er, post-translationelle modifikationer af phosphorylering, den anden er op-regulerer transcritional cofaktorer, som p33ING1, den sidste er ned-regulerer p53-hæmmere som MDM2 .
I 2000, Brodsky MH et al., undersøgt transskription mål for p53 i Drosophila. Der er beviser for, Drosophila øjne vise en svær hård sex øje fænotype under udtryk for human p53, der vil fremkalde apoptose af fantasiens øje disc celler, der forårsager tab af pigment celler, endelig at hæmme øje udvikling af Drosophila , så Drosophila kan være en model dyr for at studere funktionen af p53. Brodsky fandt ud af, at genet rpr indeholder et konsensus p53-bindingssted, der er placeret i cis-regulatorisk region af rpr, og det er også en aktivator af apoptose., Med andre beviser hævdede Brodsky, at rpr er et transkriptionelt mål for p53 . I 2001 fandt Derry og kolleger, at C. elegans ikke har et p53-gen, men indeholder faktisk et gen cep-1, der koder for proteiner, der har en lignende sekvens med protein p53. Dette C. elegans-gen koder for protein CEP-1, som har evnen til at inducere apoptose ved genotoksisk stress og er en nødvendig komponent under meiose .
i 2002 foreslog Tyner og kolleger, at p53 spiller en rolle i reguleringen af organismernes aldring., For at studere funktionen af p53, skabte de gensplejsede mus med muterede p53 ved at slette exons 1-6 og en upstream-regionen af vildtype p53-genet (p53+/+), kaldet p53+/m. Det fungere som vildtype p53 og har øget modstand mod spontane tumorer, der er bedre end vildtype p53. I eksperimentet overvågede de musene indeholdende p53+/m, p53+/+og p53+/ -. p53+ / – betyder musene, der taber en kopi af vildtype p53-genet., Resultaterne viste, at ingen af musene med p53+/m udviklet livstruende tumorer, men mere end 80% af mus med p53+/- og mere end 45% af mus med p53+/+ udviklede disse typer af tumorer. Leder indersiden af tumorer, lokaliseret tumor læsioner blev observeret i 2 ud af 35 p53+/m mus, i modsætning til forskellige tumorer som lymfomer, og osteosarcomas blev fundet i p53+/- og p53+/+ mus. Under dette forsøg observerede de også, at medianalderen på p53+/m var 96 uger, mens medianalderen på p53+/m var 116 til 118 uger., Tyner og kolleger undersøgte også muligheden for, at den kortere levetid for p53+/m var forbundet med aldring. De fandt, at efter 18 måneder begyndte p53 + /m musene at tabe sig og styrke, som for p53 + /m mus blev reducerede vægte observeret i alderen 30-36 måneder. p53 + / m mus udviser også lordokyphosis. Afhængig af røntgenanalyse udviste p53 + /m mus reduceret knogletæthed i en alder af 12 måneder, og det vil blive alvorligt i en alder af 18 måneder. Dette er et symbol på osteoporose og osteoporose er en markør for aldring hos mennesker og mus . Tyner et al., testet også tolerancen for stress, da denne evne også er en markør for aldring . De udførte 3 mm punch biopsier i ryggen af gamle og unge bedøvede p53+/m og p53+/+ mus. Deres resultater viste, at mange gamle p53+/m mus døde efter injiceret standarddosis Avertin, hvilket indikerer, at gamle p53+/m mus var mindre tolerante over for stresset .
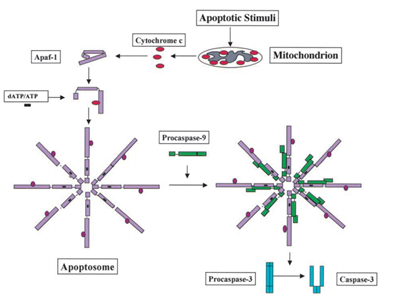
I 1991, blev det konstateret, at p53 har evnen til at fremkalde apoptose, mens der i 2003; Mihara og andre forskere fandt, at p53 også har en apoptose rolle i mitokondrierne ., Da nogle mitokondrielle proteiner har evnen til at aktivere cellulær apoptose enten ved aktive caspaser eller neutraliserende cytosoliske hæmmere. I eksemplet af cytochrom c-induceret caspase, efter at have modtaget apoptose signal, cytochrom c er frigivet fra intermembrane plads af mitokondrier, og så til gengæld binder sig til Apf-1, der eksisterer som en inaktiv monomer, inducerer dens konformationelle ændre, og øger dens bindende affinitet for dATP/ATP ved 10-fold end Apaf-1 binder dATP/ATP alene. Derefter binder den komplekse Apaf-1-cytochrom c til dATP/ATP, danner apoptosomet., Derefter rekrutterer caspase-rekrutteringsdomænet (kort) af Apaf-1 eksponeret i apoptosomet, rekrutterer procaspase-9 og derefter autoaktiverer sig selv. Det endelige kompleks spalter og aktiverer derefter andre caspaser, såsom caspase-3, som igen efterfølgende spalter vigtige molekyler i cellen, hvilket forårsager kromatinkondensation, DNA-fragmentering og til sidst fører til apoptose . Figur 2 viser cytokrom c-induceret caspase aktiveringsvej.
Mihara m Et al. forskere fandt, at vildtype p53-genet hurtigt kan translokeres til den mitokondrielle overflade af tumorceller. I eksperimentet, fandt de, at nogle stress-induceret vildtype p53 protein har evnen til at translocate til mitokondrierne thymocytter i human-eller musen celler efter apoptose på grund af DNA-skader, og hypoxi. Derefter inducerer disse vildtype p53-proteiner permeabilisering af mitokondrier og forårsager en række ændringer, der forekommer i mitokondrier som frigivelse af cytokrom c ved at danne kompleks med bcl2 og BCL .l .,
som et godt klinisk resultat med ringe bivirkning er genterapi populær. Ved udgangen af 2005 var der 1020 genterapiforsøg i databasen Journal of Gene Medicine. Blandt disse forsøg blev 66% af genterapierne udført på kræftpatienter, og 58 forsøg med dette anvendte rAd-p53, et rekombinant adenovirus, der koder for det humane p53-gen. I April 2004 blev en rekombinant human adenovirus-p53-injektion (Gendicin) lanceret formelt. Gendicin bruges til behandling af pladecellecarcinom i hoved og hals, og det blev godkendt af den statslige Food and Drug Administration i Kina den okt. 16, 2003., Det blev det første genterapiprodukt i verden, der blev godkendt af den kinesiske regering .
genet p53 blev opdaget at regulere metabolisme i 2005. For at overføre fra G1 til S-fase skal celler have tilstrækkelig råmaterialestøtte til DNA, organeller og proteinsyntese. For at regulere denne proces er nogle kontrolpunkter nødvendige. En af dem er det glukoseafhængige kontrolpunkt ved G1/S. det reguleres af den AMP-aktiverede proteinkinase (AMPK). Når glukose er opbrugt, kan AMPK phosphorylere protein p53, hvilket igen inducerer cellestopning og undgår celledød., Celler, der støder på den p53-afhængige arrestation, genindtræder cellecyklussen, når glukose er tilstrækkelig .
det er kendt, at inaktivering af p53 er nødvendig for dannelsen af tumorer. Bykov et al. VJ og Snydel EL et al. påpege, at ukorrekt funktion af p53 kan føre til spredning af en eksisterende tumor . Ventura og hans medarbejdere gjorde nogle eksperimenter for at teste denne hypotese. De gendannede funktionen af endogen p53 i primære autoktone tumorer for at undersøge konsekvensen af p53-reaktivering., Resultatet viste, at p53-reaktivering var ansvarlig for regression af autoktone tumorer. Det betyder, at inaktiveret p53-protein kan føre til tumorudvikling . Xue og andre forskere gjorde også et eksperiment for at teste konsekvensen af reaktivering af p53 på tumorer. De brugte reversibel RNA-interferens (RNAi) til at regulere ekspressionen af endogen p53 hos mus med leverkræft. I eksperimentet bruges do .ycyclin (Do.) til at genaktivere p53, da udtrykket af p53 undertrykkes fuldstændigt, når do. mangler og hurtigt gendannes, når do. tilføjes., Ved behandling med Do.blev p53 miRNA slukket, hvilket igen medfører øget ekspression af p53. Resultatet viste, at tumorerne i do.-behandlede mus bliver uopdagelige efter 12 dage, mens tumorer i ubehandlede mus voksede hurtigt. For at teste konsekvensen af forbigående reaktivering af p53 behandlede de mus med Do.i 4 dage og stoppede derefter. Resultatet viste, at selv en to-dages behandling kan forårsage regression af tumorer, og 4 dages behandling kan få tumorerne til at regressere fuldstændigt., De påpegede også, at under tumorregression kan transientreaktiveret p53 udløse cellulær senescens, ikke apoptose. Samme år fandt Hu, at embryonal implantation i p53-/- hunmus er reguleret af Leukæmihæmmende faktor (LIF). LIF er et udskilt cytokin og er vigtigt for blastocystimplantation. Genkodning LIF er identificeret som p53 målgenet og p53 bindingssted er placeret i intron 1 i både mennesker og mus .