Johdanto
toisin Kuin eläimiä, jotka voivat siirtyä välttää haitallisia ympäristö, kannattomat kasvit näytteille erittäin kehittynyt sopeutumista monimutkainen ympäristön olosuhteet. Tämän syvällisen sopeutumiskyvyn saavuttamiseksi tarvitaan solujen välistä viestintää. Solujen viestintä kasveissa liittyy vankka solunsisäinen signalointi käsittely ja monimutkainen solujen välinen signalointi verkkojen., Vasta nyt on ainakin yhdeksän signalointi aineita, nimeltään kasvi hormoneja, mukaan lukien auxin, brassinosteroid (BR), cytokinin, gibberelliinien (GA), etyleeniä, jasmonic happo (NY), strigolactone (SL), abskissihappo (ABA) ja salisyylihapon (SA) löysi (Druege et al., 2016; Verma et al., 2016). Geneettiset ja fysiologiset tutkimukset ovat paljastaneet näiden hormonien kriittiset roolit ja toimintamekanismit kasvien kasvussa ja kehityksessä (Gray, 2004)., Perustuu aiemmat tutkimukset, auxin, BR, GA, SL, ja cytokinin pääasiassa toiminto normaalin kasvien kasvua ja kehitystä, kun taas ABA, eteeni, JA, ja SA on tärkeä rooli kasvien kasvua vastaus eri bioottisten ja abioottisten korostaa (Pieterse et al., 2009; Santner ym., 2009; Denance ym., 2013). Ja myös joitakin näiden hormonien on dual roolit, esimerkiksi ABA myös näytelmiä tärkeä rooli siementen kehitys-ja lepokauden (Seo ja Koshiba, 2002)., Vaikka kukin hormoni, pelaa hallitseva rooleja tiettyjä näkökohtia, monet hormonit on päällekkäistä toimintaa ja vuorovaikutusta eri hormonien hallita monia kehityksellisiä näkökohtia, ja kasvu vastauksena endogeenisen kehityksen ja ulkoisia vihjeitä.
Auxin ja BR ovat kaksi suurta kasvua edistävien hormonien luokkaa., BR, ryhmä kasvi-erityinen steroidi hormoneja, jotka voivat vuorovaikutuksessa muiden kasvihormonit, kuten auxin, cytokinin, eteeni, GA, JA, ja SA ja säädellä monenlaisia kasvin kasvun ja kehityksen prosesseja, mukaan lukien siementen itävyys, solujen venymä, verisuonten eriyttäminen, ilmaraot muodostumista ja liikkuvuutta, kukinnan ja miehen hedelmällisyyteen (Saini ym., 2015). Mielenkiintoista on, että auxin hallitsee myös jokaista näistä prosesseista, mikä viittaa näiden kahden hormonin vuorovaikutukseen kasvien kehityksen hallitsemiseksi., Tässä tarkastelussa, me hahmotella signaalitransduktiota auxin ja BR perustuu viime edistymistä ja tarkistaa ylikuulumisen välillä auxin ja BR-välitteistä kasvien kasvua ja kehitystä.
Auxin-signalointireitti
Auxin oli ensimmäinen tunnistettu kasvi hormoni, koska sen rooli kasvi hakuisuustyypin painovoiman tai valoärsykkeisiin., Myöhemmin auxin oli kemiallisesti määritellyt indoli-3-etikkahappoa ja osoittanut pelata välttämätöntä roolia lukuisia kasvien kehitykseen ja fysiologisiin prosesseihin, mukaan lukien embryogenesis, organogeneesin aikana, verisuonten eriyttäminen, juuri ja ampua kehitystä, tropic kasvun ja hedelmien kehitys (Estelle, 2011).
geneettinen analyysi Arabidopsis, molekyyli mekanismi taustalla auxin signaalin transduktio on ollut hyvin tutkittu. LIIKENNE-ESTÄJÄ RESPONSE1 (TIR1) oli ensimmäinen tunnistettu ydinalan reseptorin ja auxin (Ruegger et al., 1998; Dharmasiri ym., 2005)., TIR1 koodaa ydinaseiden proteiinia kuuluvat F-box proteiinia kuin alayksikköön SCF E3 ubikitiinipromoottori ligase proteiini monimutkainen (Gray et al., 1999, 2002; Hellmann ym., 2003; Quint ym., 2005), lisäksi TIR1, on olemassa kolme uutta F-box proteiineja eli AUXIN SIGNALOINTI F LAATIKKO Proteiineja (AFBs), jotka osoittavat, auxin-sitova aktiivisuus ja sovitella auxin signalointi Arabidopsis (Badescu ja Napier, 2006). TIR1-reseptorilla voi olla yhteisvaikutuksia aux/IAA (auxin/indoli-3-etikkahappo) – proteiinien (Dharmasiri et al., 2003)., Aux/IAA-proteiinit ovat auxin-signaloinnin negatiivisia säätelijöitä ja Arabidopsis genomiin koodattuja aux/IAA: n jäseniä on 29. AUX/IAA proteiinit voisivat olla vuorovaikutuksessa luokan transkription sääntelyviranomaisten, auxin vastaus tekijät (ARF), välittäjänä transkription vastauksia auxin. Korkeassa auxin tasolla, AUX/IAA proteiineja vuorovaikutuksessa TIR1 kuten coreceptor ja auxin, ja voi olla ubiquitinated, jonka SCFtir1 monimutkainen ja siten olla huonontunut kautta ubikitiinipromoottori–proteasomi-reitin (Gray et al., 2001; Lanza ym., 2012)., Tuhon AUX/IAA repressors, että auxin transkription sääntelyviranomaisten ARFs, jotka sisältävät 23 jäsenyydet voidaan vapauttaa AUX/IAA sorron ja siten välittävät auxin vastauksia aktivointi tai sorron kohde-geenien (Guilfoyle ja Hagen, 2007). Eri sarjaa F-box proteiinia ja AUX/IAA tai ARFs päätellä monimutkaisuus aikana auxin signaalin transduktio (Goh et al., 2012; Guilfoyle, 2015; Salehin et al., 2015).,
koordinoitu toiminta Aux/IAA transkription repressors ja ARF transkriptio tekijät tuottaa monimutkainen geeni-sääntelyn verkostoja, jotka olivat raportoineet myös Physcomitrella (Lavy ym., 2016). Äskettäin todettiin, että CULLIN1 (CUL1) alayksikköön SCF vuorovaikutuksessa TIR1 ja siten säätelee TIR1-substraattien vakaus-ja auxin signalointi (Wang et al., 2016). Vuorovaikutusta TIR1 ja Aux/IAA vaikuttaa myös spatiaalinen rakenne Aux/IAAs, ohjataan cyclophilin isomeraasi LRT2 riisi (Jing et al., 2015)., HEAT SHOCK FACTOR 90 (HSP90) ja co-esiliina SGT1, vastaavasti, vuorovaikutuksessa TIR1 ja siten säätelee TIR1 vakautta, joka vaikuttaa vuorovaikutuksen TIR1 ja Aux/IAA ja auxin signalointi (Wang et al., 2016).
Lisäksi TIR1-riippuvainen kanoninen auxin-signalointireitti, auxin on viime aikoina raportoitu, saada erilaisia kehityksellisiä ratkaisuja kautta ei-kanoninen auxin-signalointi mekanismi., Tässä ei-kanoninen auxin tunnistus prosessi, ARF3/ETTIN ohjaa geenien ilmentyminen kautta vuorovaikutus prosessi-erityinen transkriptio tekijät, jotka erittäin rikastuttaa auxin-välitteisen kasvi kehityksen monimuotoisuutta (Simonini et al., 2016, 2017).
BR Signalointireitti
BRASSINOSTEROIDI havaittiin ensin siitepölyssä sen kyvystä edistää solujen venymistä. Myöhemmin todettiin, että BR pelaa roolit monenlaisia kasvien kasvua näkökohdat ja voi vastata bioottiset ja abioottiset korostaa., Nykyään BR-signaalin transduktio koulutusjakson oli suurelta osin selvennetty yhdistelmiä eri menetelmiä, kuten molekyyli genetiikka, biokemia, proteomiikan ja genomiikan, jne. Solu-pinta-kinaasi BRASSINOSTEROID INSENSITIVE1 (BRI1) oli tunnistettu reseptorin BR jotka voivat sitoutua solunulkoinen verkkotunnuksen BRI1 ja aktivoida sen kinaasin toimintaa ja siten kytke signaali cascade säädellä transkription (Li ja Chory, 1997; Wang et al., 2001; Kinoshita ym., 2005; Kim and Wang, 2010; Clouse, 2011; Hothorn et al., 2011; hän ym., 2011; ai ym., 2012)., Kun käsitys BR, BRI1 vuorovaikutuksessa co-reseptorin BRI1-ASSOCIATED KINASE 1 (BAK1) ja sen homolog SOMAATTISTEN EMBRYOGENESISRECEPTOR Kinaasien (SERKs) muodostaa enemmän aktiivinen BR-reseptorin monimutkainen (Li et al., 2002; Nam and Li, 2002; Wang et al., 2005; Tang et al., 2008; Gou ym., 2012). Aktivoitu BRI1 phosphorylates kaksi substraatteja plasma-kalvo-ankkuroitu reseptori-kuin sytoplasman kinaasien: BRASSINOSTEROID-SIGNALOINTI KINASES1 (BSK1) ja PERUSTAVA ERO GROWTH1 (CDG1) (Tang et al., 2008; Kim et al.,, 2011), joka puolestaan phosphorylates a PP1-tyyppi fosfataasi-nimisen BRI1-SUPPRESSOR1 (BSU1) aktivoida BSU1, mikä BSU1 dephosphorylation ja inaktivaatio, että GSK3-kaltainen kinaasi BRASSINOSTEROID INSENSITIVE2 (BIN2). Kinaasiaktiivisuuden BIN2 on myös estää histonideasetylaasi HDA6, joka on vuorovaikutuksessa ja deacetylates klo K189 ja BIN2., Kun BR tasot ovat alhaiset, BRI1 on lepotilassa, koska sen kielteisiä säädin, BRI1 KINAASI-INHIBIITTORI 1 (BKI1) ja proteiini fosfataasi 2A (PP2A), kun taas BIN2 fosforyloida kaksi BR homologisia transkriptio tekijät, BRASSINAZOLE RESISTANT1 (BZR1) ja BZR2 (myös nimeltään BES1 varten BRI1-EMS-VAIMENNIN 1) (Hän et al., 2002; Wang ym., 2002; Yin ym., 2002; Mora-Garcia ym., 2004; Kim et al., 2009, 2011; Kim ja Wang, 2010). Kun BR tasot ovat korkeat, BIN2 on inaktivoitu, ja BZR1 ja BZR2 ovat dephosphorylated, jonka PP2A, ja siirtyä tumaan muuttaa ilmaus tuhansia BR vastaus geenejä (He et al.,, 2005; Yin ym., 2005; Sun ym., 2010; Tang et al., 2011; Yu et al., 2011).
Synergiaa BR ja Auxin Signalointi
Auxin ja BR-signaalin polkuja, pelata erilaisia rooleja, mutta ne osoittivat myös, toisiaan tukevia ja toisistaan riippuvaisia vuorovaikutuksesta monenlaisia kehityksellisiä prosesseja. Esimerkiksi sekä auxin-että BR-signaalit voivat edistää solujen laajenemista ja ne voivat olla synergisesti vuorovaikutuksessa hypocotyls-venymän edistämiseksi (Nemhauser et al., 2004)., Vastaus yksi kaksi koulutusjaksojen edistäminen alkeisvarsi venymä edellyttää toimintaa muihin ja keskinäinen riippuvuus BR ja auxin väyliä (Nemhauser et al., 2004). Auxin lisääntynyt alkeisvarsi pituus villin tyypin kasveja, mutta ei BR-tunteeton mutantti bri1-116, ja tämä auxin-tunteeton fenotyyppi bri1-116 tukahdutettiin hallitseva gain-of-function mutantti bzr1-1D, mikä osoittaa, BR tai aktiivinen BZR1 tarvitaan auxin edistäminen alkeisvarsi venymä., On havaittu, että BR signalointi suppenee kanssa SUPPRESSANT PHYTOCHROME B4-3 (SOB3) hillitsemään solujen venymä ja alkeisvarsi kasvua koskevan sääntelyn kautta auxin aiheuttama PIENI AUXIN JOPA RNA19 (SAUR19) ilmaus (Favero et al., 2017). Toisaalta, sen auxin säännelty transkriptio tekijä, PIENI ELIN KOKO 1 (SMOS1) on äskettäin todettu hallita solun laajeneminen kautta suora vuorovaikutus SMOS2/DLT -, jäsen GRAS perhe transkription co-regulators, joka on myönteinen rooli BR signalointi riisi (Kim et al., 2009; Tong ym., 2012; Hirano ym., 2017)., Auxin liittyvät mutantteja, kuten iaa3 ja arf6/arf8 olivat vähemmän herkkiä BR kuin oli villin tyypin alkeisvarsi venymä, ja poistetaan yliherkkyyttä bzr1-1D auxin, mikä viittaa siihen, BR ja BZR1 edistäminen alkeisvarsi venymä edellyttää, ARF6/8. Genomin laajuinen ChIP-Seq-analyysi paljasti, että ARF6 osakkeiden suuri määrä genomista tavoitteita (noin 50%) kanssa BZR1 ja valo – /lämpötila-säänneltyjen transkriptio tekijä PIF4 CHIP-Seq analyyseissa (Oh et al., 2014)., BZR1 ja PIF4 vuorovaikutuksessa ARF6 ja aktivoida jaettu kohde-geenien sitoutuminen yhteinen tavoite geenit yhteistoiminnallisesti aikana hypocotyls venymä (Oh et al., 2014) ja monet näistä päällekkäisistä kohdegeeneistä koodaavat soluseinän proteiineja, jotka osallistuvat solujen laajenemiseen.
Brassinosteroid ja auxin myös tärkeä rooli ylläpito root apical meristem (RAM) (Durbak et al., 2012). Pässi koostuu pienestä ryhmästä harvoin jakautuvia soluja, joita kutsutaan hiljaiseksi keskukseksi (QC), jota ympäröivät kantasolut, jotka aiheuttavat erilaisia toot-kudostyyppejä., Huolto juuri kantasolujen väestöstä on säännelty WUSCHEL LIITTYVIEN MÄÄRÄYSTEN 5 (WOX5) (Sarkar et al., 2007). WOX5 rajoittuu QC: hen auxin-signaloinnin avulla ja helpottaa PLT-geenien (Aida et al., 2004; Ding ja Friml, 2010). Mutaatiot BR reseptorin geeni BRASSINOSTEROID TUNTEETON 1 (BRI1) aiheuttaa poikkeavaa solusyklin etenemisen RAM-muistia ja aiheuttaa pienemmän RAMs (Gonzalez-Garcia et al., 2011; Hacham ym., 2011). Auxinin tiedetään stimuloivan BR: n (Chung et al.,, 2011), mutta BR: n aktiivisuus ei vaikuta PIN-geenien ilmentymiseen (Hacham et al., 2011). Br-mutanttien juurikärjen fenotyypeissä ei näy samaa kuin auxinin mutanteissa(Gonzalez-Garcia et al., 2011), mikä osoittaa, että BR RAM riippumatta auxin.
Brassinosteroid ja auxin signaalit ovat myös synergisesti tarvitaan säteittäinen kuvio muodostumista verisuonten niput (Ibanes et al., 2009)., Jonka yhdistelmiä matemaattinen mallinnus ja biologisia kokeita, auxin maxima, joka on perustettu epäsymmetrinen auxin polar liikenne, mutta ei muutoksia auxin tasoilla on tärkeää, että paikannus johtosolukkokimpuista. BR-signaalin osoitettiin toimivan auxin Maximan kanssa yhdenmukaisten solujen määrän edistävänä signaalina provaskulaarisessa kehässä. Näin perustaminen määräajoin järjestely johtosolukkokimpuista ampua on alle koordinoituja toimia näiden kahden kasvi hormoneja (Ibanes et al., 2009)., Molemmat signaalit ovat myös mukana kasvi root kehitystä ja vuorovaikutusta BR ja auxin välittyy BREVIS RADIX (BRX) tämän prosessin aikana. BRX on tärkeää, että korko-rajoittaa biosynteesiä BR ja BR eksogeeninen sovellus voi pelastaa brx mutantti vikoja. Lisäksi, auxin-reagoiva geenien ilmentyminen on maailmanlaajuisesti heikentynyt vuonna brx mutantti, ja ilmaus BRX on vahvasti aiheuttama auxin ja tukahdutti BR, mikä tarkoittaa BR biosynteesiä ja auxin signalointi on yhdistetty kautta palautetta silmukka, johon BRX aikana root kehitystä (Mouchel et al., 2006).,
Brassinosteroideilla ja auxinilla on myös synergistisiä rooleja lateraalisen juurikehityksen aikana. BRs pääasiassa toiminto sivusuunnassa root primordia aloittamista, kun auxin tarvitaan sekä aloittamista ja syntyminen vaiheissa sivusuunnassa root muodostumista (Casimiro et al., 2001; Bhalerao ym., 2002; Benkova ym., 2003; Bao ym., 2004). Aikana nämä prosessit, BRs lisätä LRP aloittamista edistämällä acropetal auxin liikenteen root mutta ei vaikuttaa endogeenisen IAA-tason (Bao ym., 2004)., Kaikki nämä raportit viittaavat siihen, että ylikuulumisen välillä BR ja auxin on tärkeä rooli sääntelyn kasvien kasvua ja kehitystä.
BR Säätelee Auxin Signalointi
Lisäksi keskinäinen riippuvuus ja yhteistyö auxin ja BR signaaleja aikana kasvin kehitystä, BR voisi sovitella auxin signaalin reitti on useita tasoja. BZR1 vuorovaikutuksessa ARF proteiineja suoraan kohde-useita auxin signalointi osien ja geenien mukana auxin aineenvaihduntaa, kuten liikenne ja signalointi, mukaan lukien AUX/IAA, Nastat, TIR1, ja ARFs, jne. (Sun ym., 2010)., Todettiin, että Aux/IAA proteiinit ovat mukana BR vastauksia ja iaa7/axr2-1 ja iaa17/axr3-3 mutantit osoittivat poikkeavaa BR herkkyys ja poikkeava BR aiheuttama geenien ilmentymisen urut-riippuvaisella tavalla (Nakamura et al., 2006). Eksogeeninen brassinolidi (BL)-hoito voisi indusoida auxin-reagoivien geenien kuten IAA5: n, IAA19: n, IAA17: n jne.ilmentymistä. ja ilmaisu edellä mainitut geenit on alas-säännellään BR biosynteettiset mutantti-de-etiolated2 (det2), joka osoittaa, että toiminnallinen BR biosynteesi on osittain tarvitaan auxin-riippuvainen geenien ilmentyminen (Nakamura et al.,, 2003; Kim et al., 2006). Lisäksi BR vaikuttaa myös auxin virtausta säätelemällä ilmaus auxin viejät, kuten PIN4 ja PIN7 (Nakamura et al., 2004). Aikana kasvi gravitropism vastauksia, BRs voisi parantaa polar kertymistä auxin viejä PIN2-root meristem alueelle, ja siten vaikuttaa uudelleenjako auxin juuresta kärkeä kohti venymä-alueilla ja tulos ero IAA tasoilla sekä ylemmän ja alemman puolin juuret aiheuttaa kasvi gravitropism., Tämän prosessin aikana, BR aktivoitu ROP2 on tärkeä rooli muokkaava toiminnallinen lokalisointi PIN2-asetuksen avulla kokoonpano/kokoonpano F-actins (Li et al., 2005). Lisäksi tutkimukset osoittivat, että vähentynyt BL käsitys ja/tai pitoisuus voi aiheuttaa CYP79B2, geeni, joka koodaa entsyymiä muuntaa tryptofaania indoli-3-acetaldoxime ja siten vaikuttaa jakelu (Kim et al., 2007).
lisäksi todettiin, että BR signaali voi säädellä auxin signalointi-ulostulo sen negatiivinen säädin GSK3 kinaasi BIN2., Sen auxin vastaus tekijä ARF2 tunnistettiin BIN2 vuorovaikutuksessa proteiinin hiiva kaksi-hybridi näyttö ja kinaasi-määritys osoitti, BIN2 voi fosforyloida ARF2. Sen fosforylaatioon ARF2 johtaa menetys sen DNA: ta sitova kyky ja sorron toiminnan kohde-geenien (Vert, et al., 2008). ARF2 on bzr1-kohdegeeni ja sen ilmentymistä vähennetään BR-hoidolla (Sun et al., 2010)., Lisäksi, BIN2 voi fosforyloida ARF7 ja ARF19 tukahduttaa niiden vuorovaikutus AUX/IAAs ja siten parantaa transkription toimintaa niiden kohde-geenien SIVUSUUNNASSA URUT RAJOJA-DOMAIN16 (LBD16) ja LBD29 säännellä sivusuunnassa root organogeneesin aikana (Cho et al., 2014). Kuitenkin, BR on vain vähäinen merkitys tämän prosessin aikana, ja BIN2 on valvonnassa TRACHEARY ELEMENTTI ERIYTTÄMINEN ESTÄVÄ TEKIJÄ (TDIF)–TDIF-RESEPTORIN (TDR) – moduuli (Cho et al., 2014). Yhdessä, BR voi säädellä auxin reponses kautta vaikuttaminen eri auxin signalointi osia.,
Auxin Säätelee BR Signalointi
toisaalta, auxin voi myös säädellä BR signaalin polku tiettyjä näkökohtia. Dwarf4: n ilmentyminen, joka on tärkeä hydroksylaasi Br-biosynteesin kannalta endogeenisen BR-tason hallitsemiseksi, on auksiiniriippuvainen. Auxin-hoito voisi merkittävästi stimuloida KÄÄPIÖN4 ilmentymistä ja auxin voisi estää BZR1: n sitoutumisen KÄÄPIÖN4 promoottoriin. Induktio DWARF4, jonka auxin vaatii auxin signalointi reitin, mutta ei BR-signalointireitti (Chung et al., 2011; Yoshimitsu ym., 2011)., CPD katalysoi C-3 hapettumista BR aktivoituu BRX, oletetun transkriptio tekijä toimii alavirtaan auxin signalointi (Mouchel et al., 2006). Lisätutkimuksia riisi osoittaa, että eksogeeninen auxin voi parantaa transkriptio ilmaisun tasoa BR reseptorin geeni OsBRI1, mikä viittaa siihen, että auxin parantaa BR signalointi kautta asetus BR reseptoreihin (Sakamoto et al., 2013). Lisäksi OsBRI1: n promoottorilla on auxin-response element (AuxRE) – aihe, jonka kohteena ovat ARF-transkriptiotekijät., Lisäksi mutantti tutkimukset osoittavat, että kun mutaatio AuxRE, induktio ilmaus OsBRI1, jonka auxin on kumottu ja myös ilmaus OsBRI1 on alas säännelty arf mutantti (Sakamoto et al., 2013). Se on raportoitu, että OsARF19 sitoutuu promoottori OsBRI1 ja positiivisesti säätelee sen ilme, joka sitten aktivoi BR signalointi (Zhang et al., 2015). BES1 voi sitoutua promoottori PIENI AUXIN-UP-RNA-15 (SAUR15) ja sovitella BR varhaisen reagoinnin geeni Arabidopsis, ja tämä sitoutuminen voisi olla tehostettua by auxin hoito (Walcher ja Nemhauser, 2012)., Yhdessä auxin voi vaikuttaa myös BR-vasteisiin ja br-säännösteltyyn kasvien kasvuun ja kehitykseen.
Loppuhuomautukset ja tulevaisuudennäkymät
viimeisen lähes neljän vuosikymmenen aikana auxin-BR-reitin vuorovaikutuksia koskevat tutkimukset ovat herättäneet yhä enemmän tutkijoiden kiinnostusta. Fysiologisten, molekyylien, geneettisten ja biokemiallisten välineiden laite on suuresti syventänyt ymmärrystämme tästä asiasta., Perustuu aiemmat tutkimukset, BR ja auxin ovat mukana synergisesti useita kasvi kehitykselliset prosessit, mukaan lukien: alkeisvarsi venymä, johtosolukkokimpuista kehitystä, root kehitystä ja tropisms, jne. Keskinäinen riippuvuus ja yhteistyö auxin ja BR ovat monimutkaisia ja niihin liittyy lukuisia prosesseja molekyylitasolla, jakamalla sama kohde-geenien, säännellään toisiaan useita tasoja (Kuva 1).
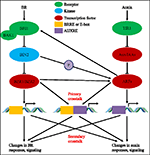
KUVA 1. Malli auxin-brassinosteroid (BR) ylikuulumisen., Arabidopsiksessa Br-ja auxin-signaalin käsitykset tunnistaa vastaavasti BRI1-ja TIR1-reseptoreista. BR sitoutuu solunulkoinen verkkotunnuksen BRI1 ja edistää se on vuorovaikutuksessa co-reseptorin BAK1 muodostaa enemmän aktiivinen BR-reseptorin monimutkainen, joka puolestaan johtaa dephosphorylation ja inaktivointi BIN2. Inaktivointi BIN2 johtaa dephosphorylation kaksi BR homologisia transkriptio tekijät BZR1 ja BZR2, jotka siirtyvät tumaan aktivoida transkriptio geenien, jotka sisältävät BRRE tai E-box niiden promoottori-alueella., BIN2 myös voi fosforyloida ARF7 ja ARF19 tukahduttaa niiden vuorovaikutus AUX/IAAs ja siten parantaa transkription toimintaa niiden kohde-geenien. TIR1 kuitti auxin signalointi ja vuorovaikutuksessa AUX/IAA proteiineja, kuten co-reseptoriin ja auxin. AUX/IAA sitten on hajonnut läpi ubikitiinipromoottori–proteasomi-reitin, ja auxin transkription sääntelyviranomaisten auxin vastaus tekijät (ARFs) ovat vapautettu AUX/IAA sorron ja aktivoida transkriptio geenien kanssa auxin reagoiva elementit (AUXRE) niiden sääntelyn alue., Jotkut ARFs voi myös sitoutuu promoottori BRI1 ja positiivisesti säätelee sen ilme, joka sitten aktivoi BR signalointi. Ensisijainen ylikuuluminen tapahtuu aktivoimalla geenejä, jotka sisältävät sekä BRRE/E-box ja AUXRE niiden promoottori-alueella, jolloin molemmat signaalinvälitysreittien suoraan säädellä transkription. Toissijainen ylikuuluminen tapahtuu kautta geenien, jotka ovat joko auxin tai BR reagoiva, mutta toimintaa, jotka ohjaavat geenien, jotka säätelevät vastaus ja signalointi muita hormoneja.,
Fosforylaation asetuksen keskeinen rooli BR signalointi koulutusjakso, erityisesti aikana käsitys prosessi, BR on koettu kautta BRI1 kinaasi-reseptorin ja BAK1 kinaasi co-reseptoreihin, ja lopulta ohjaa BR säännellä geenien ilmentyminen kautta vaikuttaa loppupään transkriptio tekijät, kuten BES1/BZR1 toimintaa (He et al., 2005; Yin ym., 2005; Sun ym., 2010; Tang et al., 2011; Yu et al., 2011). Ubikitinaatiosäätely vaikuttaa kuitenkin välttämättömältä auxin-signaloinnille., Kun auxin sitoutuu TIR1-reseptorin, joka toimii ubikitiinipromoottori E3-ligase, aktivoitu TIR1 E3-ligase ubiquitinates AUX/IAA proteiineja, johtaa hajoamista näitä repressors ja de-tukahduttaa ARF transkriptio tekijät, ja lopulta aiheuttaa auxin säännellä geenien ilmentyminen kuvio muuttuu ja kasvu vastauksia (Gray et al., 1999, 2002; Hellmann ym., 2003; Quint ym., 2005). Koska se on havaittu, että BIN2 kinaasi, joka on tunnettu toiminta BR signalointi, voi fosforyloida ja parantaa toimintaa ARFs kuten ARF2 ja ARF7 (Vert, et al., 2008; Cho et al., Vuonna 2014), on mielenkiintoista testata, jos kinaasien, kuten BIN2, jotka ovat mukana BR signalointi, voi myös vuorovaikutuksessa muiden auxin signalointi osat kuten TIR1-reseptorin tai AUX/IAA repressors, ja vaikuttaa TIR1 E3-ligase toimintaa tai AUX/IAA proteiinia stabiiliudet. Toisaalta, rooli ubiquitination BR signalointi on myös korjattava, varsinkin jos TIR1 E3-ligase voi suoraan vuorovaikutuksessa BR signalointi osia ja säännellä niiden proteiini stabiiliudet.,
lisäksi, käyttäen auxin vastaus DR5 ja muut auxin toimittajat, se on havaittu, että auxin säätelee kasvien kasvua ja kehitystä kudosten tai solujen riippuvainen tavalla. Erilaiset transcriptional tuotokset riippuen solun ja ympäristön kontekstista (Clark et al., 2014; Etshells ym., 2016; Lavy ym., 2016)., Vaikka spatiotemporaalisia BR signalointi on osoitettu control juurikasvua läpi vihamielinen toiminta auxin (Chaiwanon ja Wang, 2015), se on vielä tuntematon, jos kudosten tai solujen BR signalointi, joka voisi olla visualisoitu pBZR1:BZR1-YFP, on myös tärkeää ohjata muita prosesseja kuin root kehitystä. Lisäksi sukupolven yksityiskohtainen kudoksen tai kellarissa kartta auxin ja BR jakaumat on tällä hetkellä mahdollista käyttäen fluoresenssi-aktivoida solun lajittelu tai laser microdissection yhdessä korkean resoluution geenien ilmentyminen analyysi., Tämä johtaa lopulta osoite, jos auxin ristitalks kanssa BR kudoksen tai solun tavalla.
Kirjailijasuoritukset
kaikki kirjoittajat osallistuivat tämän arvostelukäsikirjoituksen kirjoittamiseen. Lisätietoja siitä, mikä on tekijyys, katso tekijän ohjeita.,
Rahoitus
Tämä työ tukee avustuksia National Natural Science Foundation of China (Hankkeet 31500229 ja 31770305), Shandongin Maakunnassa Natural Science Foundation of Major Basic Research Program (2017C03), Shandongin Maakunnan Varoja Erottaa Nuoret Tutkijat (2014JQ201408).
eturistiriita Lausunto
kirjoittajat ilmoittavat, että tutkimus on tehty ilman mitään kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdollisia eturistiriitoja.
Clouse, S. D. (2011)., Brassinosteroidisignaalin transduktio: reseptorikinaasin aktivaatiosta kasvien kehitystä sääteleviin transcriptionaaliverkkoihin. Kasvisolu 23, 1219-1230. doi: 10.1105 / tpc.111.084475
PubMed Abstrakti | CrossRef Koko Teksti | Google Scholar
Estelle, M. (2011). Auxin signalointi: synteesistä Järjestelmäbiologiaan; Aihekokoelma kylmän kevään satamasta näkökulmia biologiaan. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Google Scholar
Guilfoyle, T. J. (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar