Johdanto
Nasu kuolleisuus on yksi suurimmista valinnan piirteitä sikatalouteen ja vaikuttavat emakkojen, porsaiden, ja ympäristö. Siksi porsaiden kuolleisuus on monimutkainen fenotyyppi ja riippuu kapasiteetti kylvää nostaa sen jälkeläisiä, mutta on myös toiminto, syntymäpaino -, johto -, ja valinta (Knol ym., 2002)., Kuitenkin myös geenin resessiivinen vikoja edistää porsaiden kuolleisuutta, vaikka vain muutamia esimerkkejä on raportoitu aiemmin (Murgiano et al., 2012; Matika ym., 2019). Niissäkin tapauksissa, joissa mutaation vaikutus on vakava, tehokas valinta tällaista mutaatiota vastaan vaikeutuu vähäisen taajuuden vuoksi. Monissa vakavia vikoja, tsygootit kuolla hyvin varhain raskauden, jättämättä jälkeäkään muut kuin puuttuminen homotsygootit väestön suuri (Derks et al., 2019).,
Sisäsiitoksen vaikutuksia kaupallinen sikoja on yleensä pidettävä kurissa valikoiva jalostukseen laski kuolleisuutta porsaille parantamalla sekä äidillisiä kykyjä ja nasu elinkelpoisuuden (Olijslagers, 2018). Kuitenkin, vaihtoehdot taustalla resessiivinen geenin viat eivät ole hyvin kiinni sisällä jalostukseen arvot, ja mahdollisesti drift korkeammat taajuudet seurauksena voimakas valinta (Georges ym., 2019). Lisäksi nämä muunnokset voidaan säilyttää myös tasapainotettaessa valintaa heterotsygoottisessa tilassa olevan korreloivan positiivisen vaikutuksen (Derks et al., 2018).,
resessiiviset viat vain marginaalisesti vaikuttavat porsaiden kokonaiskuolleisuuteen (Alonso-Spilsbury et al., 2007). Porsaiden kuolleisuuteen vaikuttavat muunnokset ovat kuitenkin erittäin tärkeitä, koska nämä muunnokset vaikuttavat suoraan tuotantoon ja eläinten hyvinvointiin (Baxter ym., 2013; Rutherford ym., 2013). Kuitenkin, eläinten väestön hallintaa, matalien taajuuksien esiintyminen vikoja on yleensä huonosti dokumentoitu (usein hyvin yleisesti käytetään), ja oireyhtymät ovat usein vain tunnustettu kun ne ovat saavuttaneet korkean taajuuden., Tämä on erityisen tärkeää oireyhtymissä, jotka eivät johda hyvin erillisiin fenotyyppeihin. Siksi, vaikka kaupallinen pesivät pikku-seuranta voidaan tehdä erityisiä oireyhtymät ja tehokkaasti valitsemalla vastaan erityisiä matalan taajuuden oireyhtymät siksi tarvitaan uusia lähestymistapoja.
tässä työssä kuvaamme löytö erittäin heikentävä oireyhtymä kaupallinen sika väestö kautta tutkimuksen perusteella yhdistetty medium-tiheys SNP-taulukot ja koko-genomin sekvensointi (WGS)., Tutkimus johti 16-bp frameshift-poiston tunnistamiseen SPTBN4-geenissä, ja homotsygooteissa ennustettiin selkeitä fenotyyppisiä seurauksia. Kantoaallon taajuus on noin 9% väestöstä tutkitaan, vaikuttaa noin 0.81% väestöstä pentueet. Taajuus oli riittävän alhainen, jotta se olisi tuntematon on geneettinen perusta, ja jopa tehokkaasti tunnistamattomia tiettyyn oireyhtymä ollenkaan. Tutkimuksen toteutumisen yhteydessä yksi tiineenä ollut emakko tunnistettiin kantajasikojen siittämäksi., Vaikuttaa porsaat kärsivät myopatia ja pysty kävelemään, yleensä johtaa kuolemaan muutaman tunnin kuluttua syntymästä, täysin sopusoinnussa ennusti patologian verrattuna samanlainen ihmisen ja hiiren tapauksissa.
aineisto ja Menetelmät
Eläimet, Genotyypit, ja Pre-Processing
aineisto koostuu 31,839 eläimiä synteettinen villisika line kanssa suuri valkoinen tausta. Linjaa ylläpidetään ja kasvatetaan Topigs Norsvin nucleus-tiloilla, ensisijaisesti tuotannon ja terveysominaisuuksien valinnassa., Eläimet genotyypitettiin Illumina GeneSeek custom 50K SNP-sirulla (Lincoln, NE, USA). Eläimet, joiden genotyypit puuttuvat > 0, 15 poistettiin. Me heitetään pois merkit, jotka eivät täytä seuraavia suodatusehtoja: vähintään puheluiden hinta 0,85, pieni alleelia taajuus > 0.01, ja Hardy-Weinberg mittasuhteet tarkka testi p-arvo alle P < 10-12. Lisäksi markkereita tuntematon sijainti Sscrofa11.1 genomin rakentaa hylättiin, jättäen 41,573 markkereita suodatuksen jälkeen., Kaikki vaiheet suoritettiin Plink v1.90b3.30 (Purcell ym., 2007).
Haplotyyppi Vaiheistus ja Tunnistaminen SSC6 Haplotyyppi
Me tehdään haplotyyppi toteuttaminen ja kohdentaminen puuttuu sivustoja Beagle5.0 parametrin tehokkaan populaation kokoa 100, muut asetukset olivat oletusarvoissa (Browning ym., 2018). Odotetut homotsygootit arvioitiin haplotyypin taajuuden perusteella Hardy-Weinberg-periaatteen mukaisesti. Tarkkaa binomitestiä käytettiin Havaittujen homotsygoottien määrän testaamiseen odotettujen homotsygoottien määrällä., Se haplotyyppi pidettiin merkittävästi köyhdytettyä, jos P < 5 × 10-3.
Fenotyyppisiä Vaikutuksia, jotka Liittyvät SSC6 Haplotyyppi
tutkimme SSC6 haplotyyppi tietueiden kokonaismäärä syntynyt, useita kuolleena, muumioitunut porsaiden, porsimisen selviytymisen, ja imetyksen aikana selviytymisen (survival noin 21 päivän ikäisiä) yhteensä 9,666 pentueet. Listasimme nämä fenotyypit kaikille CXC: lle ja cxn pentueille tunnistetut. Käytimme Welchin t-testiä arvioidaksemme, eroavatko CxC-pentueiden fenotyypit merkittävästi CxN-pentueista. P-arvo < 0.,05: tä pidettiin merkittävänä.
Koko-Genomin Sekvensointi Analyysi ja Ehdokas Variantti Tunnistaminen
aineisto koostuu 71 koko genomi sekvensoitiin yksilöitä väestö tutkimuksen kohteena. Kaikki 71 näytteet olivat myös läsnä meidän aineisto 31,839 eläimiä, joiden genotyyppi oli 50K. 71 näytteiden kokonaistilavuus 1,93 Tbp (tera base paria) alkaen 14.16 miljardia 150-bp pariksi-end lukee (Taulukko S3). Näytteet sekvensoitiin Illumina HiSeq 2000-kanavalla. Suuntasimme sekvenssejä, että Sscrofa11.1 genomin rakentaa käyttäen BWA-MEM versio 0.7.,15 (Li ja Durbin, 2009), keskimääräinen mappability 98,9% ja otoksen kattavuus vaihtelee 8,8-14,8 X (keskiarvo 10,9 X). Samblasteria käytettiin PCR-kaksoiskappaleiden poistamiseen (Faust and Hall, 2014). Samtoolsia käytettiin tiedostojen lajitteluun,yhdistämiseen ja indeksointiin (Li et al., 2009). Kartoitus ja laatutilastot syntyivät Qualimapin (Okonechnikov et al., 2016). Variantti soittamalla suoritettiin Freebayes v1.1.0 seuraavat asetukset: –min-base-laatu 10 –min-vaihtoehtoinen-osa 0.2 –haplotyyppi-pituus 0 –min-vaihtoehtoinen-count 2 (Garrison ja Marth, 2012)., Variantteja Phred laatu pisteet < 20 hylättiin (Li et al., 2009). Variantteja selitettiin käyttäen Ensembl variant effect predictor (VEP, release 96) (Mclaren et al., 2016). Missense-varianttien vaikutus ennustettiin käyttämällä lajittelu-intoleranttia siedettävästä (SIFT) (Kumar et al., 2009). LD-analyysi suoritettiin Plink v1.90b3.30: llä (Purcell ym., 2007) seuraavilla asetuksilla –chr-set 18, –r2, ld-ikkuna-r2 0,8.
SPTBN4-proteiinin linjaus
proteiinin linjaus villin tyypin ja mutanttiproteiinin välillä suoritettiin Clustalolla (Madeira et al.,, 2019) ja visualisoitiin ESPript 3: lla (Robert and Gouet, 2014). Edelleen visualisointi ja validointi suoritettiin käyttäen JBrowse genome viewer versio 1.12.1(Skinner et al., 2009).
Validointi Syy 16 bp SPTBN4 Poistaminen
PCR tehtiin käyttäen 60 ng genomista DNA: ta, 0,4 µm kunkin pohjamaali, 1.8 mM MgCl2, ja 25 yksikköä/ml OneTaq® DNA-Polymeraasi (OneTaq® 2X Master Mix Vakio Puskuri, New England Biolabs) valmistaja on PCR-puskuri lopullinen määrä on 12 µl., Alkuperäinen denaturointi 1 min 95°C seurasi 35 sykliä 95°C 30 s 55°C 45 s, 72°C 90 s, jonka jälkeen 5 min pidennys 72°C. PCR-alukkeet SPTBN4 ovat TCAAGGGTGCAGGCTCTTTC eteenpäin ja GGTAGGAAGCTCGAAGTGGG taaksepäin. Eteenpäin pohjamaali oli väriaine-merkitty joko 6-FAM tuottaa fluoresoivasti merkitty PCR-tuotteen havaittavissa on ABI 3730 DNA-tutkimus (Applied Biosystems). Fragmenttikoot määritettiin ABI: n GeneMapper software 5: llä.,
Histopatologinen Tutkimus
Kaksi vaikuttaa porsaiden alle 1 viikon ikäisenä lähettää patologian osasto Royal Animal Health (Deventer) tutkittavaksi. Makroskooppisesti kaikki havainnot olivat normaalirajoilla. Luuston lihaksen etujalan, selkä-lihas-ja takapuolella jalka sekä eläimiä otettiin näyte rutiini H&E värjäys ja PTAH värjäystä. Lihaskudosta säilytettiin erillisissä purkeissa ja se kiinnitettiin formaldehydiliuokseen 4% puskuroituna (=formaliiniliuos 10%, puskuroitu)., Sen jälkeen, kudos oli upotettu parafiini ja viipaloitu 2 µm: n mukaan standard operation procedure (SOP RAH). Sen jälkeen, diat olivat deparaffinized ja rutiininomaisesti värjättiin hematoksyliini-ja eosiini (H&E) automaattinen värin kone. Samanaikaisesti uusia dioja 2 µm lihaskudoksen sekä positiivinen kontrolli liu ’ uta lihaskudoksen olivat valmiita manuaalinen värjäys ”phosphotungstic acid hematoksyliinillä,” lyhennettynä PTAH. Tätä värjäystä suositellaan luustolihasten poikkijuovien osoittamiseksi.,
Kasvatus Arvot ja Yhdistyksen Analyysi
tässä tutkimuksessa olemme arvioineet 63 piirteitä käytetään jalostukseen ohjelma. Kunkin tutkittavan ominaisuuden vastemuuttujana käytettiin poistettuja arvioituja jalostusarvoja (DEBV). Kaikkien arvioitujen ominaisuuksien arvioitu jalostusarvo (EBV) purettiin Garrick et al: n kuvaamalla menetelmällä. (2009). EBV jokainen eläin oli saatu rutiinia geneettinen arviointi kaupallinen kasvatus-ohjelma (Topigs Norsvin) käyttämällä eläin-malli., Se luotettavuuksien eläintä varten deregression poimittiin geneettinen arviointi perustuvat menetelmät Tason ja Meyer (2004). Myös sääntelyn purkamiseen käytetyt heritabiliteetit on poimittu rutiininomaisesta geneettisestä arvioinnista. Myös DEBV: n arvioituun luotettavuuteen perustuvat painotuskertoimet arvioitiin Garrick et al: n mukaan. (2009) käyttää arvoa 0,5 skalaari c. Varmistaa laadun DEBV, vain eläimiä, joiden painokerroin on suurempi kuin nolla ja luotettavuus DEBV suurempi kuin 0.,Yhdistysanalyyseissä käytettiin 20. Debv: n luotettavuus saatiin myös Garrick et al: n mukaan. (2009).
Assosiaatioanalyysit tehtiin asreml-ohjelmalla (Gilmour et al.,, 2009) soveltaen seuraavia lineaarinen sekoitettu eläinten malli:
missä DEBVij on havaittu DEBV eläimen j, w on painokerroin jäljellä, µ on yleinen DEBV tarkoita väestöstä, Ri on operaattorin asema (lasken haitallinen alleeli) 4 mutaatio i, aj on additiivinen geneettinen vaikutus arvioitu käyttäen sukutaulu-pohjainen keskimääräinen suhde, matriisi, ja jäljelle jäävä virhe. Yhdistykset, joiden a-log10 (p-arvo) on suurempi kuin viisi, julistettiin merkittäviksi.
Tulokset
1.,5 Mb Segmentin Kromosomi 6 Vaikuttaa Imetyksen Selviytymisen Sioilla
Me analysoidaan 31,638 eläimiä yhdestä puhdasrotuisia villisikoja line (synteettinen line kanssa suuri valkoinen tausta), joiden genotyyppi on Sian 50K SNP-siru (Sscrofa11.1 build) (Warr ym., 2019). Analyysi paljasti, 1,5 Mb segmentin kromosomi 6 (SSC6:48.75–50.25) osoittaa alijäämää homotsygoottisiksi liittyy vähentää imetyksen survival (Taulukot 1 ja 2). Se haplotyyppi on erottelevat kohtuullisella alleelia taajuus 4,5%: iin (9,0% carrier frequency) väestön tutkimus., Haplotyypin esiintymistiheys on vaihdellut viimeisen vuosikymmenen aikana, mutta vähentynyt viimeisten 3 vuoden aikana (Kuva S1). Testasimme, johtuiko taajuus heterotsygoottisesta etuefektistä. Löysimme kuitenkin enimmäkseen negatiivisesti tärkeä valinta piirteitä lukuun ottamatta lanne syvyys ja tiineyden pituus (Taulukko 3), mikä viittaa siihen, että taajuus on puhtaasti seurausta geneettinen ajautuminen.

Taulukko 1 SSC6 haplotyyppi ominaisuudet.,

Taulukko 2 Carrier-by-kantaja pentueet ssa 24%: n lasku imetyksen selviytymisen verrattuna carrier-ei-operaattorin pentueet. Merkittävät tulokset on lihavoitu.

Taulukko 3 Piirteet merkittävästi liittyvät heterotsygootteja kantajia SPTBN4 poistetaan.
52 carrier-by-kantaja (CxC) pentuetta ei näytä merkittävästi vähentää kokonaismäärä syntynyt elävänä tai eläimiä., Kuitenkin, imetys selviytyminen on vähentynyt noin 24% CxC-pentueet verrattuna carrier-by-noncarrier (cx series cxn) astutukset, mikä osoittaa, että homotsygoottinen porsaita kuolee imetyksen aikana (Taulukko 2). Seuraavaksi tarkastelimme CXC pentueiden huomioita ajasta ja kuolinsyystä. Tämä paljasti, että useimmat porsaat, jotka kuolivat ensimmäisten 24 tunnin aikana syntymän jälkeen. Suurin osa noista porsaista oli maanviljelijöiden mukaan ”heikkoa porsasta syntyessään.,”
Koko-Genomin Sekvensointi Analyysi Paljastaa, 16-bp Frameshift Poistetaan SPTBN4 kuin Todennäköisesti Aiheuttava Variantti
tunnistaa syy-mutaatio, me tutkittiin koko perimän sekvenssi tietoja 71 eläimiä perusjoukosta tutkitaan ja tunnistetaan viisi harjoittaja eläimiä. Sidos epätasapainoa (LD) analyysi paljasti, 267 SNP ja indel variantit korkea LD (r2 > 0.8) kanssa SSC6 haplotyyppi (Taulukko S1), suurin osa on täydellinen LD (247 vaihtoehtoja)., Vain viisi vaihtoehtoa mahdollisesti vaikuttaa koodaavan sekvenssin (kolme näin, yksi frameshift, yksi liitos-tunnustaja). Kolme näin variantteja ennustetaan olevan siedetty SEULOA (pisteet > 0.18, Taulukko S1), kun taas splice-tunnustaja variantti vaikuttaa geeni koodaa 28 bp peptidi tuntematon toiminto, todennäköisesti syy. Kuitenkin, yksi variantti täysin LD (r2 = 1) haplotyyppi oli ennustettu, on suuri vaikutus; 16-bp frameshift poistetaan eksonin 26 SPTBN4 gene (6:g.48801280delGACGGTGTACGCCGGT) (Kuvat 1A, B)., Se frameshift poistaminen (ENSSSCP00000031537:s.Arg1902fs) esittelee 30 romaani aminohappoja ja ennenaikaisen lopetuskodonin, tuottaa heikentynyt ja typistetty spectrin beta ei-erythrocytic 4-proteiinin (SPTBN4). Mutantteja ei ole lopullinen 662 aminohappoja villityypin proteiinia (Kuva 1C), mukaan lukien pleckstrin homology (PH) domain tarvitaan proteiinia liikenteen kalvot (Wang et al., 2018). Se SPTBN4 proteiini on jäsenenä beta-spectrin proteiineja ja on aktiini, joka yhdistää solukalvon aktiini tukirankansa., SPTBN4 mutaatioita häiritä cytoskeletal koneiden valvontaan oikea lokalisoinnin ioni-kanavia myelinated hermoja aiheuttaen moottorin neuropatiat (Parkinson ym., 2001; Wang ym., 2018).

Kuva 1 (A) SPTBN4 gene malli. Sairastuneen 26. eksonin sijainti on merkitty punaisella. B) kuvituskuva 16-bp: n poistosta. Kuvassa villityyppi ja mutanttieksoni. (C) mutantin (Mt) ja villin tyypin (WT) SPTBN4-proteiinijakson kohdistaminen. Mutaatio indusoi 30 uutta aminohappoa ja ennenaikaisen stop-kodonin.,
Genotyypin Viisi CxC-Pentueet Vahvistaa SPTBN4 Poistetaan niin Todennäköisesti Syyllinen
Me, joiden genotyyppi oli viisi CxC-pentueet 16-bp poistaminen, joka oli ainakin kaksi porsaat (vaihteluväli 2-6), joka kuoli ensimmäisten 48 h kuluttua syntymästä. Nämä viisi poikuetta tuottivat 53 porsasta, joista 19 oli homotsygootteja 16 bp: n poistoon (Taulukko 4). Kaikki 19 homotsygoottista porsasta kuolivat 48 tunnin sisällä syntymän jälkeen (18 24 tunnin sisällä). 34 loput porsaat (8 villi tyyppi, ja 26 lentoyhtiötä), vain 1 kuoli 48 h, todennäköisesti aiheuttamat muut (ympäristö) tekijät.,

Taulukko 4 Genotyypin todennäköinen syy-16-bp SPTBN4 frameshift poistetaan viidessä harjoittaja-by-kantaja pentueet. Summa per genotyyppi on lihavoitu.
Porsaat ovat Homotsygoottisia SPTBN4 Poistetaan Kärsivät Myopatia ja Takajalan Halvaantuminen
Olemme seurata yksi viime CxC-pentue (porsimista päivämäärä: 28. huhtikuuta 2019), joka tuotti kuusi tervettä, kaksi vaikuttaa (näytteet: 9912, 9916) (Kuva 2A), ja kolmesta kuolleena syntyneet porsaat., Vahvistimme homotsygoottisen SPTBN4-poistotilan kahden sairastuneen porsaan osalta (taulukko S2). Lisäksi havaitsimme terveillä yksilöillä neljä heterotsygoottista kantajaa ja kaksi homotsygoottista villityyppistä porsasta. Yksi kuolleena syntyneistä porsaista (näyte: 9921) oli myös homotsygoottinen poistoa varten, kun taas kaksi muuta olivat heterotsygoottisia. Vaikuttaa porsaat kärsivät äärimmäinen lihasten heikkous (Kuvat 2B, C), halvaantuminen takajalkojen raajojen, ja vapinaa (S1-Video). Siksi porsaat eivät kyenneet kävelemään eivätkä juomaan.,
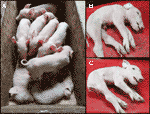
Kuva 2 (A) Kaksi vaikuttaa porsaiden (elossa) yhdessä kuusi terveet sisarukset. Porsaat ovat peräisin yhdestä CXC: n parittelusta, joka oli farrowedissa 28.huhtikuuta 2019. B) sairastunut urospossu 9912. C) sairastunut naaraspossu 9916.
Vaikuttaa Porsaiden Puute Rajat Juovia Selkä-ja Takajalan Luuston Lihaksia
Histopatologinen tutkimus paljasti hajallaan rappeutumista lihas kuituja sekä porsaiden, ja fokaalisesti kuolio ja vaskuliitti selkä lihas yksi porsaat (ID = 9912)., Lisäksi phosphotungstic acid hematoksyliinillä (PTAH) värjäys osoittaa, poikkeava väritys luuston lihassyiden, mikä osoittaa laskua rajat juovia, erityisesti lihaksia, selkä ja takajalat vaikuttaa eläinten (Kuva 3B), kun etujalat näyttävät ennallaan (Kuva 3A). Lasku rajat juovia on merkitty epänormaali väritys ja yleinen menetys määrä lihassyiden (Kuva 3B). Histopatologisesti havaitut muutokset takajaloissa ja selkälihaksissa viittaavat lihasdystrofiaan.,

Kuva 3 (A) poikkipinta-näkymä luuston lihaksen edessä jalka. Musta nuoli osoittaa lihassyiden normaalia väritystä (pimeää), mikä viittaa poikkijuovien esiintymiseen. PTAH Bar = 50 µm. B) luustolihaksen poikkileikkaus takajalasta. Musta nuoli osoittaa lihassyiden epänormaalia väritystä (vaaleanpunaista), mikä viittaa poikkijuovien puuttumiseen. Keltainen nuoli osoittaa normaalin värityksen ja ristijuovien läsnäolon. PTAH Bar = 50 µm.,
Keskustelua
tässä työssä kerromme romaani synnynnäinen vika aiheuttaa porsaiden kuolleisuus johtui todennäköisesti 16 bp frameshift poistetaan vuonna SPTBN4 geeni. Porsaat kärsivät äärimmäisestä lihasheikkoudesta (myopatiasta) ja kuolevat muutaman tunnin kuluessa syntymästä. Poiston odotetaan aiheuttavan spektribeetan, ei-erytrosyyttisen 4-proteiinin, täydellisen toimintakyvyn menetyksen. SPTBN4 on perheenjäsen spectrin geenit ja tarvitaan ioni-kanava klusterointi klo ranvierin kuroumia, jotka vaikuttavat aktiopotentiaalin (Deveaux, 2010)., Mutaatiot häiritä cytoskeletal koneisto, joka ohjaa oikea lokalisoinnin ioni-kanavia ja toiminta axonal verkkotunnukset pääasiassa axon alustavat segmentit (AIS) ja ranvierin kuroumia (Wang et al., 2018). Tarkemmin sanottuna vaikuttaa C-terminaali verkkotunnuksen SPTBN4 on ratkaiseva KCNQ2 kanava ihmiskaupan ja excitability klo ranvierin kuroumia (Deveaux, 2010).
jatkotoimia tutkimus-tunnistaa ihmisen ja hiiren tapauksissa todettiin, että seuraava oireyhtymä ei todennäköisesti olisi olla välittömästi tappava, mutta pikemminkin antaa vakavia myopatia., Keskipitkän tiheys SNP genotyypin tietoja, saatavilla kaikille eläinten jalostukseen väestön (N = 31,839), kantajia voi olla tunnistettavissa. Joukossa kantajia oli emakko, joka oli noin puolivälissä raskauden aikaan tunnistaminen, isä villisika, joka oli myös harjoittaja. Siitostilalle ilmoitettiin, että pentue dokumentoidaan syntyessään. Havaittu fenotyyppi vaikuttaa porsaiden (myopatia, takajalan halvaantuminen, vapinaa) oli täysin yhtenevä sen kanssa, mitä oli havaittu ihmisen potilaat, joilla on homotsygoottinen loss-of-function-tai yhdiste heterotsygoottiset mutaatiot SPTBN4 geeni (OMIM: 606214)., Kahdella ihmispotilaasta on pH-alueella funktiomutaatioita (Wang et al., 2018), joka tukee sitä, että PH-alueen menettäminen sioilla johtaisi todennäköisesti SPTBN4-proteiinin täydelliseen toimintakyvyttömyyteen. Ihmisellä samankaltaiset mutaatiot johtavat vaikeaan synnynnäiseen myopatiaan, joka johtuu lihastyypin I kuitujen, neuropatian ja kuurouden puuttumisesta (Knierim et al., 2017; Wang ym., 2018). Wang ym. (2018) myös havaittu motorinen aksonaalinen neuropatia useille potilaille ominaista synnynnäinen hypotonia, syvällinen heikkous, ja menetys syvä jännerefleksien varhain lapsuudessa., Lisäksi hermo koepaloja paljasti vähentää solmukohtien Na+ – kanavat ja ei solmukohtien KCNQ2 K+ – kanavia, paljastaen molekyyli-patologian aiheuttaa hermoston toimintahäiriöitä. Näin ollen, voimme päätellä, että tämä frameshift variantti on todennäköisesti kausaalinen mutaatio johtaa havaittu fenotyyppi ja ehtyminen homotsygootti genotyyppi väestöstä. Tulevissa tutkimuksissa voitaisiin keskittyä sian SPTBN4-geenin in vivo-tyrmäykseen, jotta syndroomaa ja siihen liittyvää fenotyyppiä voitaisiin tutkia tarkemmin.,
Emme noudata rappeutumista lihassyiden edessä jalat, kun selkä-ja takajalan lihas kuidut olivat selvästi vaikuttaa. Tämä havainto voisi osittain selittää takaraajan halvaantumisen, kun taas etujalat eivät vaikuta. Ero etu-ja takajalkojen lihasten kuituja on myös kuvattu vapisevan hiiret, jossa SPTBN4 tappio-off-toiminto mutaatioiden aiheuttaa motorinen neuropatia, takajalan halvaantuminen, vapinaa, ja keski-kuurous (Parkinson ym., 2001; Komada ja Soriano, 2002). Parkinson ym., (2001) kuvaavat vähentää hermo-johtuminen nopeudet iskias hermot hiirten kanssa vapisevan alleeleja aiheuttaa perifeerinen takajalan neuropatia. Ilmaus SPTBN4 hiirillä on rajoitettu aivojen, selkäytimen, ja iskias hermot ja ei havaittu luuston lihaskudosta, joten tämä tauti on pääasiassa hermosolujen vika. Kaiken kaikkiaan on epäselvää, mikä mekanismi aiheuttaa oireiden puuttumisen eturaajoissa. Tämä ”luonnollinen tyrmäys” sioissa voi olla hyödyllinen resurssi tutkimaan ihmisen sairauksia, kuten siat ovat yleensä parempi malli tutkimaan ihmisen sairauksia verrattuna jyrsijä., Lisäksi SPTBN4-funktion häviämisen seurauksia voidaan tutkia tarkemmin.
tutkimuksen kohteena olevan rodun efektiivisen populaation koon (ne) arvioidaan olevan noin 100 (Hidalgo et al., 2016). Eläinjalostuksessa matala Ne lisää riskiä, että haitalliset alleelit yleistyvät sattumalta. Lisäksi aiemmat tutkimukset ovat osoittaneet, että väistyvä tappava alleeleja voidaan ajaa edulliset vaikutukset heterozygotes (Derks et al., 2018; Matika ym., 2019). Matika ym.,, 2019 löytynyt resessiivinen stop-sai mutaatio MSTN geeni liittyy merkittävä kasvu lihasten syvyys heterozygotes. Emme kuitenkaan löydä tutkimuksestamme mitään todisteita mistään heterotsygoottisesta edusta. Nykyinen genomista tekniikoita, voimme nyt tunnistaa haitallisia alleeleja ajautuminen korkeampia taajuuksia, ja seurata syntymistä romaani haitallisia alleeleja tarkasti, mikä lisää tehokasta huuhtelua. Lisäksi tämäntyyppisen tutkimuksen tulos parantaa huomattavasti tietoisuutta ”piilotetuista” geneettisistä vioista sekä kasvattaja-että viljelijätasolla., Ilman ennakkotietoja harvinaiset synnynnäiset viat kirjataan usein ” heikkona porsaana.”Ja ilman, että erityisiä oireyhtymiä enää erotellaan toisistaan, jatkotoimet eivät ole mahdollisia. Useimmissa tapauksissa ei tiedetä, onko geeniperimää tai onko muita hämmentäviä vaikutuksia. Ennen genomista tietoa, oireyhtymä voidaan tunnistaa, verrattuna muissa tapauksissa, ja harjoittajat tunnistaa, mikä käytännöllisiä tietoja.
Nasukuolleisuudella on suuri taloudellinen ja eläinten hyvinvoinnin merkitys., Näin ollen löytö SPTBN4 mutaatio on johtanut välitön täytäntöönpano jalostukseen ohjelma minimoida taajuus carrier-by-kantaja astutukset. Tämä mahdollistaa välttää syntymän vaikuttaa yksilöiden, mikä parantaa eläinten hyvinvointia ja vähentää taloudellisia menetyksiä.
Johtopäätös
tässä tutkimuksessa raportoimme romaani synnynnäinen vika todennäköisesti aiheuttama väistyvä frameshift poistetaan vuonna SPTBN4 geeni sikoja. Havainnot tukevat silmiinpistävää yhtäläisyyksiä SPTBN4 liittyvät syndromic fenotyyppeihin ihmisillä ja hiirillä., Tutkimuksen avulla voidaan seurata ja puhdistaa herkästi leviävää alleelia väestöstä. Carrier-by-kantaja ristit voi olla esti, että se on esteenä vaikuttaa yksilöiden, mikä vähentää taloudellisia tappioita, ja parantaa eläinten hyvinvointia. Lopuksi, nämä ”luonnollinen aihiot” saatu jalostukseen teollisuus voi tarjota mallina ihmisten sairauksien ja lisätä ymmärrystä geenin funktiota nisäkkään haaran, ja tarjota mahdollisia malli ihmisen tauti.,
Tietojen Saatavuus Lausunto
Etiikka-Selvitys
Eettistä arviointia ja hyväksyntää ei vaadita, että eläinten tutkimus, koska tietoja käytetään tässä tutkimuksessa on saatu osana rutiini tietojen keräämistä Topigs Norsvin jalostukseen ohjelmia, ja ei nimenomaan tämän hankkeen tarkoituksena. Siksi eettisen komitean hyväksyminen ei ollut pakollista. Näytteiden keruu ja Tietojen tallennus toteutettiin tiukasti eläinten suojelusta ja hyvinvoinnista annetun Alankomaiden lain (Gezondheids – en welzijnswet voor dieren) mukaisesti., Omistajat saivat kirjallisen suostumuksen eläinten osallistumiseen tähän tutkimukseen.
Tekijäosuudet
MG, H-JM ja MD suunnittelivat ja suunnittelivat tutkimuksen. BH vastasi yleisestä järjestämisestä ja yhteydenpidosta Topigs Norsvinin ja maanviljelijöiden kanssa. MD ja ML tekivät data-analyysin. BD ja KL tekivät laboratoriotöitä. TP-V teki patologisen analyysin. MD kirjoitti käsikirjoituksen. H-JM, MG, BH, SG-V, BD, KL ja ML esittivät hyödyllisiä kommentteja ja ehdotuksia ja auttoivat käsikirjoituksen laatimisessa. Fenotyyppitiedot analysoitiin ML: lla., Kaikki kirjoittajat lukivat ja hyväksyivät lopullisen käsikirjoituksen.
Rahoitus
Tämän tutkimuksen on rahoittanut STW-Breed4Food Kumppanuutta, hankkeen numero 14283: Alkaen järjestyksessä fenotyyppi: havaita haitallinen vaihtelu, jonka ennustus toimintoja. Tämä tutkimus tukee taloudellisesti NWO-TTW ja Breed4Food kumppanit Cobb-Euroopassa, CRV, Hendrix Genetics ja Topigs Norsvin. Lisäksi tätä tutkimusta tuki IMAGE-hanke (Horisontti 2020, nro 677353). Rahoittajilla ei ollut mitään roolia tutkimuksen suunnittelussa, tiedonkeruussa ja analysoinnissa, julkaisupäätöksessä eikä käsikirjoituksen valmistelussa., HPC-klusterin käytön mahdollisti Katagrofood (Shared Research Facilities Wageningen UR).
Disclaimer
tietoja käytetään tässä tutkimuksessa on saatu osana rutiini tietojen keräämistä Topigs Norsvin jalostukseen ohjelmia, ja ei nimenomaan tämän hankkeen tarkoituksena. Siksi eettisen komitean hyväksyminen ei ollut pakollista. Näytteiden keruu ja Tietojen tallennus toteutettiin tiukasti eläinten suojelusta ja hyvinvoinnista annetun Alankomaiden lain (Gezondheids – en welzijnswet voor dieren) mukaisesti.,
eturistiriita
ML ja BH ovat työntekijät Topigs Norsvin Research Center, research institute läheisesti yksi rahoittajien (Topigs Norsvin).
Kaikki jäljellä olevat kirjoittajat julistaa, että tutkimus on tehty ilman mitään kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdollisia eturistiriitoja.
muut Breed4Food partners Cobb Europe, CRV, Hendrix Genetics, ilmoittavat, ettei ole kilpailevia intressejä tähän tutkimukseen.,
Kiitokset
kirjoittajat kiittää Frank van der haaren ja Toon Janssen järjestämisestä viestinnän tilan. Kiitämme Manon Houbenia hänen panoksestaan patologisiin tutkimuksiin. Kiitämme Mout Viller ottaa video ja kuvat vaikuttaa ja terve porsaiden syntymän jälkeen. Kiitämme Jürgen Harliziusta siitä, että hän huolehti porsaista luovutuksen jälkeen. Kiitämme Gerda van Eldikiä näytteenotosta. Kiitämme Egbert Knolia ja Egiel Hanenbergia Topigs-Norsvinin yleisestä tuesta ja ohjauksesta.,
oheismateriaali
Video S1 | Video osoittaa sekä vaikuttaa yksilöiden syntymän jälkeen.
Taulukko S1 | Genomista vaihtelut korkea LD kanssa SSC6 haplotyyppi.
taulukko S2 / CXC-pentueen genotyypin määritys, mukaan lukien kaksi sairastunutta yksilöä (porsituspäivä: 28.huhtikuuta 2019). Taulukossa on esitetty homotsygoottisen del/del-tilan kahden sairastuneen porsaan osalta 99129916.
taulukko S3 / WGS-näytteiden kartoitus-ja kattavuustilastot.
Kuva S1 | SPTBN4 poistaminen harjoittaja taajuuden 2007-2019.