소개
과는 달리는 동물 이동할 수 있는 부작용을 피하기 위해 환경을 착 식물을 전시 높게 개발에 적응한 복잡한 환경 조건에 따른 것입니다. 이 심오한 적응력을 달성하기 위해서는 세포 간의 통신이 필요합니다. 식물에서의 세포 대 세포 통신은 강력한 세포 내 신호 처리 및 복잡한 세포 간 신호 네트워크를 포함한다., 지금까지 있는 적어도 구호물질,이라는 식물 호르몬 등 auxin,brassinosteroid(BR),시토키닌,gibberellins(GA),에틸렌,jasmonic acid(JA),strigolactone(SL),abscisic 산(ABA),그리고 살리실산(SA)발견했(Druege et al.,2016;Verma 외., 2016). 유전 적 및 생리 학적 연구는 식물 성장 및 발달에서 이들 위의 호르몬의 중요한 역할과 기능적 메커니즘을 밝혀냈다(Gray,2004)., 기반으로 이전 연구 auxin,BR,GA,SL 및 시토키닌 주로 기능 일반적인 식물의 성장과 발전하는 동안,ABA,에틸렌,JA,과 SA 의 중요한 역할을 식물 성장에 대응하는 다양한 biotic and abiotic(Pieterse et al.,2009;산트 너 외.,2009;Denance 외., 2013). 또한 이러한 호르몬 중 일부는 이중 역할을하며,예를 들어 ABA 는 종자 개발 및 휴면기에 중요한 역할을합니다(Seo and Koshiba,2002)., 지만 각 호르몬 놀이 주된 역할을에서의 특정 측면을 겹쳐 많은 호르몬 활동과 상호 작용의 다른 호르몬 제어 많은 발달 측면과 성장에 대응하여 생 개발 및 외인성습니다.
옥신과 BR 은 성장 촉진 호르몬의 두 가지 주요 부류입니다., BR,그룹의 공장 특정 스테로이드 호르몬을 수 있는 상호 작용으로 다른 모종은 식물 호르몬과 같은 auxin,시토키닌,에틸렌,GA,JA,사고 조절하는 다양한 식물의 성장과 발달 과정을 포함한 종자 발아,셀 신장,혈관 분화,기공이 형성 및 운동,꽃과 남성의 불임(Saini et al., 2015). 흥미롭게도,이러한 각 프로세스에 의해 제어되는 또한 옥신,제안이 두 호르몬 작용을 제어하는 식물의 개발이다., 이 검토에서,우리는 것입니다 개요 신호 전달의 auxin 및 BR 기반에서 최근 진행하고 검토 사이 누화 auxin 및 BR 중재 식물의 성장과 개발.
Auxin 신호 통로
Auxin 처음으로 인식되었으로 식물 호르몬에서의 역할 때문에 식물을 현상을 중력 또는 가벼운 자극., 나중에 auxin 었 화학적으로 식별로 indole-3-아세트산과에 필수적인 역할을 과다는 식물의 개발 및 생리 프로세스,including embryogenesis,organogenesis,혈관 분화,루트 및 촬영을 개발,트로픽의 성장,그리고 과일을 개발(에스텔,2011).
Arabidopsis 에서 유전자 분석을 사용하여 옥신 신호 전달의 기초가되는 분자 메커니즘을 잘 조사했습니다. 수송 억제제 RESPONSE1(TIR1)은 옥신의 첫 번째 확인 된 핵 수용체였다(Ruegger et al.,1998;Dharmasiri 외., 2005)., TIR1 은 F-box 단백질에 속하는 핵 단백질을 SCF E3 유비퀴틴 리가 제 단백질 복합체의 서브 유닛으로 코딩한다(Gray et al.,1999,2002;헬만 외.,2003;퀸트 외. 2005 년)외에 TIR1,세 추가 F-box proteins 즉 AUXIN F 신호 상자에 단백질(AFBs)을 보여주는 auxin-바인딩 작업 중재 auxin 신호 Arabidopsis(Badescu 및 Napier,2006). TIR1 수용체는 AUX/IAA(옥신/인돌-3-아세트산)단백질 그룹과 상호 작용할 수있다(Dharmasiri et al., 2003)., AUX/IAA 단백질은 auxin 신호 전달의 음성 조절 자이며 Arabidopsis 게놈에 인코딩 된 aux/IAA 의 29 개 구성원이 있습니다. AUX/IAA 단백질은 전사 조절제 인 옥신 반응 인자(arf)의 부류와 상호 작용하여 옥신에 대한 전사 반응을 매개 할 수 있습니다. 에서 높은 auxin 수준,AUX/IAA 단백질 상호 작용 TIR1 로러의 auxin,과될 수 있는 실 ubiquitinated 에 의해 SCFtir1 복잡하고 따라서 저하될을 통해 ubiquitin–proteasome pathway(회색 et al.,2001;란자 외., 2012)., 의 파괴에 AUX/IAA repressors,auxin transcriptional 레귤레이터 ARFs 을 포함하 23 회원이 될 수 있습에서 발표 AUX/IAA 억제하고 따라서 중재 auxin 반응 활성화에 의해 또는 억압의 대상 유전자(길포일 및 하겐,2007). F-box 단백질과 AUX/IAA 또는 ARFs 의 다른 세트는 옥신 신호 전달 동안 복잡성을 추론합니다(Goh et al.,2012;Guilfoyle,2015;Salehin 외., 2015).,
Aux/IAA transcriptional repressors 와 ARF 전사 인자의 조정 된 작용은 Physcomitrella 에서도보고 된 복잡한 유전자 조절 네트워크를 생성한다(Lavy et al., 2016). 최근에,SCF 의 CULLIN1(CUL1)서브 유닛이 TIR1 과 상호 작용하여 TIR1 기질 안정성 및 옥신 시그널링을 조절한다는 것이 발견되었다(Wang et al., 2016). TIR1 과 Aux/IAA 사이의 상호 작용은 또한 rice 의 cyclophilin isomerase LRT2 에 의해 제어되는 Aux/IAAs 의 공간적 구성에 의해 영향을 받는다(Jing et al., 2015)., 열 충격 요인 90(HSP90)와 co-chaperone SGT1,각각의 상호 작용하 TIR1 며 따라서 규제 TIR1 안정성에 영향을 미치는 간의 상호 작용 TIR1 및 Aux/IAA 및 auxin 신호(왕 et al., 2016).
외 TIR1 의존하는 표준 auxin 신호 경로,auxin 는 최근에 보고되었을 이끌어내는 다양한 범위의 개발을 통해 비정규 auxin 신호 메커니즘이 있습니다., 이 비 정규 auxin 감지 프로세스,ARF3/곰곰이트 유전자 발현을 통해 상호작용 과정-특정 전사 인 요소,는 매우 풍요롭게 auxin 중재 식물 다양성을 발달(Simonini et al., 2016, 2017).
BR 신호 전달 경로
BRASSINOSTEROID 는 세포 신장을 촉진하는 능력에 대해 꽃가루에서 처음 발견되었습니다. 나중에 BR 은 광범위한 식물 성장 측면에서 역할을하며 생체 및 비 생물 스트레스에 반응 할 수 있음이 밝혀졌습니다., 요즘 BR 신호 전달 경로는 크게 명확히 하여 조합의 다른 방법을 포함하여 분자 유전학,생화학,단백질,유전체학,등등. 세포 표면 니 BRASSINOSTEROID INSENSITIVE1(BRI1)었으로 확인되는 수용체의 BR 할 수 있는 바인딩하여 세포의 도메인 BRI1 및 활성화 니 활동하고 따라서 전환에서 신호를 캐스케이드을 조절하는 전사(리튬 및 Chory,1997;왕 et al.,2001;키노시타 외.,2005;Kim and Wang,2010;Clouse,2011;Hothorn 외.,2011;그녀 외.,2011;오 외., 2012)., 에 따라 인식의 BR,BRI1 와 상호 작용 co-receptor BRI1 관련 KINASE1(BAK1)및 그 체 체세포 EMBRYOGENESISRECEPTOR KINASEs(SERKs)을 더 활성 BR receptor complex(리 et al.,2002;Nam and Li,2002;Wang et al.,2005;당나라 외.,2008;고우 외., 2012). 활성화 된 BRI1 은 혈장 막 고정 수용체 유사 세포질 키나아제의 두 가지 기질을 인산화시킨다:BRASSINOSTEROID-SIGNALING KINASES1(BSK1)및 구성 차등 성장 1(CDG1)(Tang et al.,2008;김 외.,,2011),차례로 BRI1-SUPPRESSOR1(bsu1)이라는 PP1 형 포스 파타 아제를 인산화시켜 BSU1 을 활성화시켜 BSU1 탈 인산화 및 gsk3 유사 키나제 브라 시노 스테로이드 INSENSITIVE2(BIN2)를 불 활성화시킨다. BIN2 의 키나아제 활성은 또한 BIN2 의 K189 에서 상호 작용하고 탈 아세틸 화되는 히스톤 탈 아세틸 화 효소 HDA6 에 의해 억제된다., 때 BR 수준이 낮은,BRI1 은 무부하로 인해 부정적인 레귤레이터,BRI1 키니아제 억제물 1(BKI1)과 단백질 인산 2A(PP2A),동 BIN2phosphorylate 두 BR 동종 transcription factors,BRASSINAZOLE RESISTANT1(BZR1)및 BZR2(이라고도 BES1 에 대한 BRI1-EMS 억제 1)(그 et al.,2002;왕 외.,2002;음과 외.,2002;Mora-Garcia 외.,2004;김 외.,2009,2011;김과 왕,2010). BR 수준이 높으면 BIN2 가 불 활성화되고 bzr1 과 BZR2 는 pp2a 에 의해 탈 인산화되고 핵으로 이동하여 수천 개의 BR 반응 유전자의 발현을 변화시킨다(He et al.,,2005;음과 외.,2005;Sun 외.,2010;당나라 외.,2011;유 외., 2011).
사이의 시너지 효과 BR 및 Auxin 신호
Auxin 및 BR 신호 경로 다양한 플레이 역할,그러나,그들은 또한 시너지 효과와 상호 의존적인 상호 작용의 넓은 범위에서 개발 프로세스입니다. 예를 들어,옥신 및 BR 신호 모두 세포 확장을 촉진 할 수 있고 상승적으로 상호 작용하여 하이포 코틸 신장을 촉진 할 수있다(Nemhauser et al., 2004)., 응답의 두 가지 중 하나로 추진에 어린 줄기 연신율이 필요합의 기능이 다른 사이의 상호 의존성을 BR 및 auxin 경로(Nemhauser et al., 2004). Auxin 증가한 어린 줄기는 길이 야생형 식물이지만 BR 을 구분하지 않는 돌연변이 bri1-116,이 auxin 을 구분하지 않는 형의 bri1-116 억제되었으로 지배적 인 기능 이득 돌연변이 bzr1-1D 을 나타내는,BR 또는 활성 BZR1 에 필요한 옥신의 승진 어린 줄기 신장., 그것은 발견되는 BR 신호로 수렴의 억압 PHYTOCHROME B4-3(SOB3)하 제어 세포 신장 및 어린 줄기 성장을 통해 규제 auxin 유도 작은 AUXIN 최대 RNA19(SAUR19)현(Favero et al., 2017). 다른 한편으로,auxin regulated transcription factor 작은 기관 1(SMOS1)최근에 발견하 제어 세포의 확장을 통해 직접적인 상호 작용과 함께 SMOS2/DLT,회원의 그라 가족의 transcriptional co-레귤레이터는 긍정적인 역할을에서 BR 에서 신호 쌀(Kim et al.,2009;통 외.,2012;히라노 외., 2017)., Auxin 관련 돌연변이와 같은 iaa3 및 arf6/arf8 덜 민감하 BR 했던 것보다 야생 유형에 대한 어린 줄기 연신 율,폐지하고 과민 반응의 bzr1-1D auxin 제안,BR 및 BZR1 승진의 어린 줄기 연신율이 필요합 ARF6/8 입니다. 게놈 차원의 칩-Seq 분석은 ARF6 이 BZR1 과 칩-Seq 분석에 의한 빛/온도 조절 전사 인자 PIF4 와 방대한 수의 게놈 표적(약 50%)을 공유한다는 것을 밝혀냈다(Oh et al., 2014)., BZR1 과 PIF4 는 hypocotyls 신장 동안 협력 적으로 공유 된 표적 유전자에 결합함으로써 ARF6 과 상호 작용하고 공유 된 표적 유전자를 활성화시킨다(Oh et al.,2014)및 이들 겹치는 표적 유전자의 대부분은 세포 확장에 관여하는 세포벽 단백질을 암호화합니다.
Brassinosteroid 와 auxin 은 또한 뿌리 정점 meristem(RAM)의 유지에 중요한 역할을합니다(Durbak et al., 2012). RAM 로 구성되어 있는 작은 그룹에 드물게 나누어로 알려져 있는 세포는 대기 전류 센터(QC),에 의해 둘러싸여 줄기 세포는 다양한 나팔 소리 조직이 유형이 있다., 뿌리 줄기 세포 집단의 유지는 WUSCHEL 관련 HOMEOBOX5(WOX5)에 의해 조절된다(Sarkar et al., 2007). WOX5 는 옥신 신호 전달에 의해 QC 로 제한되고 PLT 유전자의 적절한 발현을 촉진한다(Aida et al.,2004;딩과 프림,2010). BR 수용체 유전자 BRASSINOSTEROID INSENSITIVE1(BRI1)의 돌연변이는 램에서 비정상적인 세포주기 진행을 초래하고 더 작은 램을 유발한다(Gonzalez-Garcia et al.,2011;Hacham 외., 2011). 옥신은 BR 의 생합성을 자극하는 것으로 알려져있다(Chung et al.,,2011),그러나 BR 의 활성은 핀 유전자의 발현에 영향을 미치지 않는다(Hacham et al., 2011). Br 돌연변이 체의 뿌리 팁 표현형은 옥신 돌연변이 체와 동일하게 보이지 않는다(Gonzalez-Garcia et al.,2011),auxin 과 독립적으로 RAM 에 BR 이 작용 함을 나타냅니다.
브라 시노 스테로이드 및 옥신 신호는 또한 혈관 다발의 방사형 패턴 형성에서 상승적으로 요구된다(Ibanes et al., 2009)., 에 의해 조합의 수학적 모델링과 생물학적 실험,auxin maxima,설립하여 비대칭 polar auxin 운송,하지만 변경에 auxin 수준은 중요한 위치를 조정하는 혈관들. BR 신호는 옥신 맥시마와 일치하는 프로 혈관 고리의 세포 수에 대한 촉진 신호 역할을하는 것으로 나타났다. 따라서 촬영에서 혈관 다발의 주기적 배열의 확립은이 두 식물 호르몬의 조정 된 작용하에있다(Ibanes et al., 2009)., 두 신호는 또한 식물 뿌리 발달에 관여하며 br 과 옥신의 상호 작용은이 과정에서 BREVIS RADIX(BRX)에 의해 매개됩니다. BRX 는 br 의 속도 제한 생합성에 중요하며 BR 외인성 적용은 brx 돌연변이 결함을 구출 할 수 있습니다. 또한,auxin 반응의 유전자 발현 세계적으로 장애인에 경우 돌연변이,그리고 표현의 경우 강력에 의해 유도 된 auxin 및 억제 BR,뜻 BR 생 합성 및 auxin 신호를 통해 연결되어있 피드백 루프를 포함하는 경우 동안 루트 개발(Mouchel et al., 2006).,
Brassinosteroids 와 auxin 은 또한 측면 뿌리 발달 동안 상승 작용 역할을한다. BRs 는 주로 측면 뿌리 primordia 개시에서 기능하는 반면,옥신은 측면 뿌리 형성의 개시 및 출현 단계 모두에 요구된다(Casimiro et al.,2001;Bhalerao 외.,2002;벤코바 외.,2003;바오 외., 2004). 이러한 과정에서 BRs 는 뿌리에서 아크로 페탈 옥신 수송을 촉진시킴으로써 LRP 개시를 증가 시키지만 내인성 IAA 수준에 영향을주지는 않는다(Bao et al., 2004)., 이 모든 보고서는 BR 과 옥신 사이의 누화가 식물 성장 및 발달 조절에 중요한 역할을한다는 것을 시사한다.
BR 조절 Auxin 신호
외에 상호 의존하고 협력의 auxin 및 BR 신호는 동안 식물의 개발,BR 중재할 수 있는 auxin 신호 경로에서 여러 수준. BZR1 상호작용으로 ARF 단백질을 직접 대상에는 여러 auxin 시그널링 구성 요소와 관련된 유전자에 auxin 와 같은 물질 대사는 운송 및 시그널링 포함하여,AUX/IAA,핀,TIR1 및 ARFs,etc. (일 외., 2010)., 그것이었다는 것을 발견 Aux/IAA 단백질에 참여하고 있 BR 응답 iaa7/axr2-1iaa17/axr3-3 돌연변이 탈 BR 감도와 탈 BR-유도 유전자 발현에서는 기관에 의존 방식(Nakamura et al., 2006). 외인성 brassinolide(BL)처리는 iaa5,IAA19,IAA17 등과 같은 옥신 반응 유전자의 발현을 유도 할 수있다.,그리고 표현의 위의 유전자 다운 규제에 BR 생합성 돌연변이 de-etiolated2(det2)음을 나타내는 기능 BR 생합성는 부분적으로 필요한 옥신 의존하는 유전자 발현(Nakamura et al.,,2003;김 외., 2006). 또한,BR 은 또한 pin4 및 PIN7 과 같은 옥신 수출자의 발현을 조절함으로써 옥신 흐름에 영향을 미친다(Nakamura et al., 2004). 안 식물 gravitropism 응답이 BRs 을 향상시킬 수 있는 극의 축적 auxin 수출 PIN2 루트에 분열 조직 영역을 따라 영향을 미치의 재배포 auxin 루트에서 끝을 향해 신장 영역에서 결과의 차이를 낼 수준 모두에서 상부 및 하부 측의 뿌리를 유도하는 식물 gravitropism., 이 과정에서 br 활성화 된 ROP2 는 F-액틴의 조립/재 조립의 조절을 통해 PIN2 의 기능적 국소화를 조절하는 데 중요한 역할을한다(Li et al., 2005). 더 많은 연구가 보이는 감소 BL 인식 및/또는 농도를 유도할 수 있 CYP79B2,유전자 인코딩하는 효소 변환 트립토판을 indole-3-acetaldoxime 따라서 영향 분배(Kim et al., 2007).
또한,br 신호는 음성 조절제 GSK3 키나아제 BIN2 에 의해 옥신 신호 출력을 조절할 수 있음을 발견했다., Auxin 응답 요소 ARF2 로 확인되었다 BIN2 단백질 상호 작용에 효모 두-하이브리드 스크린과 키 분석 결과를 보였 BIN2 수 phosphorylate ARF2. ARF2 의 인산화는 dna 결합 능력의 손실과 표적 유전자의 억압 활성을 초래한다(Vert et al., 2008). ARF2 는 bzr1 표적 유전자이며 그 발현은 BR 처리에 의해 감소된다(Sun et al., 2010)., 또한,BIN2phosphorylate 수 있습니다 ARF7 및 ARF19 을 억제하는 그들의 상호작용으로 AUX/IAAs 함으로써 향상 transcriptional 활동에서 대상 유전자의 측면 기관의 경계-DOMAIN16(LBD16)및 LBD29 을 조절하는 측면 루트 organogenesis(Cho et al., 2014). 그러나 BR 은이 과정에서 사소한 역할을하며 BIN2 는 기관 요소 분화 억제 인자(TDIF)–TDIF 수용체(TDR)모듈(Cho et al., 2014). 함께,BR 은 상이한 옥신 신호 성분에 영향을 미치는 것을 통해 옥신 레폰 스를 조절할 수있다.,
옥신은 BR 시그널링을 조절한다
반면에 옥신은 또한 특정 양상에서 BR 신호 경로를 조절할 수있다. 내인성 BR 수준을 제어하기 위해 BR 생합성에 중요한 히드 록 실라 제 인 DWARF4 의 발현은 옥신 의존적이다. 옥신 처리는 dwarf4 의 발현을 눈에 띄게 자극 할 수 있고 옥신은 DWARF4 의 프로모터에 BZR1 의 결합을 억제 할 수있다. 옥신에 의한 DWARF4 의 유도는 옥신 신호 전달 경로를 필요로하지만 BR 신호 전달 경로는 필요하지 않다(Chung et al.,2011;요시미츠 외., 2011)., BR 의 C-3 산화를 촉매하는 CPD 는 옥신 시그널링의 하류에서 작용하는 putative transcription factor 인 BRX 에 의해 활성화되었다(Mouchel et al., 2006). 더 많은 연구에서 쌀을 나타내는 외생 auxin 을 강화할 수 있는 전사 표현 레벨의 BR receptor gene OsBRI1 제안 auxin 향상 BR 을 통해 신호의 규정 BR 수용체(사카모토 et al., 2013). 또한,OsBRI1 의 프로모터는 ARF 전사 인자에 의해 표적화되는 상류 옥신-반응 요소(AuxRE)모티프를 보유한다., 또한,돌연변이 연구에의 돌연변이 AuxRE,의 유도의 식 OsBRI1 여 auxin 은 폐지되도의 식 OsBRI1 다운 규제에 arf 돌연변이(사카모토 et al., 2013). OsARF19 는 OsBRI1 의 프로모터에 결합하고 그 다음 BR 신호를 활성화시키는 발현을 긍정적으로 조절한다고보고되었다(Zhang et al., 2015). BES1 묶을 수 있습 프로모터의 작은 AUXIN-UP RNA15(SAUR15)및 중재 BR 초기 대응의 유전자에 Arabidopsis,그리고 이 바인딩에 의해 향상 될 수 있습 auxin 치료(Walcher 및 Nemhauser,2012)., 함께 복용하면 옥신은 BR 반응 및 br 조절 식물 성장 및 발달에도 영향을 줄 수 있습니다.
발언을 체결하고 미래의 관점
과거 동안의 연구에 auxin BR 통로의 상호 작용이 있을 끌었고 더 많은 연구원을 제공하고자 합니다. 생리 학적,분자 적,유전 적 및 생화학 적 도구의기구는이 문제에 대한 우리의 이해를 크게 심화 시켰습니다., 기반으로 이전의 연구,BR 및 auxin 참여하는 공동성으로 여러 식물 개발 프로세스 포함:어린 줄기 신장,혈관들 개발,루트를 개발하고 tropisms,etc. 상호 의존하고 협력의 auxin 및 BR 은 복잡하고 포함한 수많은 프로세스에서 분자 수준을 공유함으로써,동일한 대상 유전자 조절 각각 다른 상호간에 여러 수준의(그림 1).
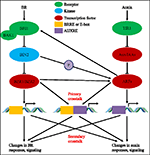
그림 1. 옥신-브라 시노 스테로이드(BR)누화의 모델., Arabidopsis 에서 BR 및 옥신 신호의 인식은 각각 BRI1 및 TIR1 수용체에 의해 인식됩니다. BR 은 BRI1 의 세포 외 도메인에 결합하고 공동 수용체 BAK1 과 상호 작용하여 bin2 의 탈 인산화 및 불 활성화를 유도하는보다 활성 BR 수용체 복합체를 형성하도록 촉진한다. 비활성화 BIN2 으로 이어질 dephosphorylation 의 두 BR 동종사 요인 BZR1 및 BZR2,이동으로 핵을 활성화하는 유전자의 전사를 포함하는 BRRE 또는 이란에서 자신의 발기인 지역이다., BIN2 는 또한 ARF7 및 ARF19 를 인산화하여 aux/IAAs 와의 상호 작용을 억제하여 표적 유전자에 대한 전사 활성을 향상시킬 수있다. TIR1 은 옥신 신호를 수신하고 옥신의 공동 수용체로서 AUX/IAA 단백질과 상호 작용한다. AUX/IAA 그는 저를 통해 ubiquitin–proteasome pathway,그리고 auxin transcriptional 레귤레이터 auxin 응답 요인(ARFs)의 발표에서 AUX/IAA 억제 활성화 유전자의 전사과 auxin 응답소(AUXRE)에서 자신의 규제 지역이다., 일부 ARFs 는 또한 BRI1 의 프로모터에 결합하여 그 발현을 긍정적으로 조절하여 br 신호를 활성화시킬 수 있습니다. 기본 크로스 토크에 의해 발생한 차등적 유전 자 활성화 현상을 모두 포함하는 BRRE/E-상자 및 AUXRE 에서 자신의 발기인이 지역 수 있도록 모두 신호 경로를 직접 조절을 전사. 보조 누가 발생합을 통한 유전자의 표현은 하나 auxin BR 또는 반응하지만,의 활동을 제어하는 유전자의 발현을 조절 하는 응답 신호의 기타 호르몬.,
인 산화 규제에서 중요한 역할을 BR 신호 경로,특히 인식 과정,BR 을 통해 인식되 BRI1receptor kinase 및 BAK1 니 co-수용체,그리고 결국은 컨트롤 BR 규제를 통해 유전자 발현을 좌우하는 다운스트림 전송 등의 요인 BES1/BZR1 활동(그 et al.,2005;음과 외.,2005;Sun 외.,2010;당나라 외.,2011;유 외., 2011). 그러나,유비퀴틴 화 조절은 옥신 신호 전달에 필수적인 것으로 보인다., 한 auxin 묶 TIR1 수용체 역할을 하는 E3ubiquitin-리가,활성 TIR1E3-리가 ubiquitinates AUX/IAA 단백질,지도 저하로 이러한 repressors and de-누르는 멍멍사 요인,그리고 결국 원인 auxin 규제되는 유전자 발현의 패턴의 변화와 성장 응답(회색 et al.,1999,2002;헬만 외.,2003;퀸트 외., 2005). BR 신호 전달에서 잘 알려진 기능을하는 BIN2 키나아제가 ARF2 및 ARF7 과 같은 ARFs 의 활성을 인산화하고 향상시킬 수 있다는 것이 발견 되었기 때문에(Vert et al.,2008;조 외., 2014 년),그것은 흥미로운 일이 될 것이를 테스트하는 경우 kinases 등 BIN2 에 관여하는 BR 신호를 수도용 기타 옥신 신호와 같은 구성요소 TIR1 수용체 또는 AUX/IAA repressors,와 영향력을 TIR1E3-리가 활동 또는 AUX/IAA 단백질 안정성. 다른 한편으로의 역할 ubiquitination 에 BR 신호 또한 해결 될 필요가 있는 경우에 특히 TIR1E3-리가의 수와 직접 상호 작용 BR 신호 구성요소와 조절이 그들의 단백질 안정성.,
이외에,사용 auxin 응답 DR5 및 기타 auxin,기자들이 관찰 되었습니다 auxin 을 조절한 식물의 성장과 발전에 조직 또는 세포에 따른 방식으로. 세포 및 환경 적 맥락에 따라 다양한 전사 출력(Clark et al.,2014;Etchells 외.,2016;라비 외., 2016)., 하지만 spatiotemporal BR 신호가 표시되어 뿌리 성장을 통해 대립과 행동 auxin(Chaiwanon 과 왕,2015),그것은 여전히 알 수없는 경우에는 조직 또는 세포 BR 신호,시각화할 수 있으로 pBZR1:BZR1-YFP,는 것도 중요하 제어 다른 프로세스 외에,루트 개발. 또한,세대의 자세한 조직이나 지하실의 지도 auxin 및 BR 배포판은 현재 사용하여 가능한 형광 활성화 셀 정렬하거나 인 레이저 조합에서 높은 해상도의 유전자 발현 분석합니다., 이것은 결국 auxin 이 조직 또는 세포 방식으로 BR 을 가진 crosstalks 경우 주소로 연결됩니다.
저자 기고
모든 저자는이 검토 원고의 작성에 참여했다. 저자를 구성하는 것에 대한 자세한 내용은 저자 지침을 참조하십시오.,
자금 지원
이 작품이 지원되는 보조금에서 National Natural Science Foundation of China(프로젝트 31500229 및 31770305),Shandong Province 자연 과학 재단의 주요 기본적인 연구 프로그램(2017C03),산동성 자금 특별한 젊은 학자들(2014JQ201408).
이해의 충돌 문
저자가 선언하는 연구가 수행되었의 부재에서 어떠한 상업 또는 금융 서비스를 제공하는 것으로 해석될 수 있는 잠재적인 이해의 충돌.나는 이것이 내가 할 수있는 유일한 방법이라고 생각한다., 브라 시노 스테로이드 신호 전달:수용체 키나아제 활성화에서 식물 발달을 조절하는 전사 네트워크에 이르기까지. 식물 세포 23,1219-1230. doi:10.1105/tpc.111.084475
PubMed Abstract|CrossRef 전체 텍스트|Google Scholar
Estelle,M.(2011). 옥신 신호:합성에서 시스템 생물학에 이르기까지;생물학의 콜드 스프링 하버 관점에서 주제 수집. 콜드 스프링 하버,뉴욕:콜드 스프링 하버 실험실 보도.나는 이것이 내가 할 수있는 유일한 방법이라고 생각한다., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar