Innledning
i Motsetning til dyr som kan bevege seg for å unngå de negative omgivelser, sessile planter viser en høyt utviklet tilpasning til de kompliserte forholdene. For å oppnå dette dyp tilpasning, kommunikasjon mellom celler nødvendig. Celle-til-celle kommunikasjon i planter innebærer robust intracellulære signal processing og intrikate intercellulær signaliserer nettverk., Til nå er det minst ni signaliserer stoffer, kalt anlegg hormoner, inkludert auxin, brassinosteroid (BR), cytokinin, gibberellins (GA), etylen, jasmonic syre (JA), strigolactone (SL), abscisic syre (ABA), og salicylic acid (SA) oppdaget (Druege et al., 2016; Graham et al., 2016). Den genetiske og fysiologiske studier har avslørt viktige roller og funksjonelle mekanismer av disse ovenfor hormoner i plantevekst og utvikling (Grå, 2004)., Basert på tidligere studier, auxin, BR, GA, SL, og cytokinin hovedsakelig funksjon under anleggets normale vekst og utvikling, mens ABA, etylen, JA, og SA spille viktige roller i å plante vekst svar på ulike biotiske og abiotiske stressfaktorer (Pieterse et al., 2009; Santner et al., 2009; Denance et al., 2013). Og også noen av disse hormonene har doble roller, for eksempel, ABA spiller også viktige roller i frø utvikling og dvalen (Seo og Koshiba, 2002)., Selv om hvert hormon som spiller en dominerende rolle i visse aspekter, mange hormoner har overlappende aktiviteter og interaksjoner av ulike hormoner styre mange utviklingsmessige aspekter, og veksten i respons til endogene utviklingsmessige og eksogene signaler.
Auxin og BR er to store klasser av vekst-fremme hormoner., BR, en gruppe av plante-spesifikke steroid hormon som kunne samhandle med andre phytohormoner som auxin, cytokinin, etylen, GA, JA, og SA, og regulere et bredt utvalg av plante vekst og utviklingsmessige prosesser, inkludert frø spiring, celle tøyelighet, vaskulær differensiering, stomata dannelse og bevegelse, blomstrende og mannlig fruktbarhet (Saini et al., 2015). Det er interessant at hver av disse prosessene er også kontrollert av auxin, noe som tyder på at disse to hormonene samspillet til å styre anlegget utvikling., I denne gjennomgangen vil vi skissere signal transduksjon av auxin og BR basert på de nylige fremskritt og gjennomgå krysstale mellom auxin og BR mediert plante vekst og utvikling.
Auxin Signaliserer Sti
Auxin ble først anerkjent som anlegg hormon som på grunn av sin rolle i anlegget tropism gravitasjonen eller lys stimuli., Senere auxin var kjemisk identifisert som indole-3-eddiksyre og som vises til å spille viktige roller i overflod av anlegget utviklingsmessige og fysiologiske prosesser, inkludert embryogenesis, organogenesis, vaskulær differensiering, rot og skyte utvikling, tropic vekst, frukt og utvikling (Estelle, 2011).
ved Hjelp av genetiske analyser i Arabidopsis, den molekylære mekanismen bak den auxin signal transduksjon har blitt godt undersøkt. TRANSPORT-HEMMER RESPONSE1 (TIR1) var den første som identifiserte kjernefysisk reseptor av auxin (Ruegger et al., 1998; Dharmasiri et al., 2005)., TIR1 blir en kjernefysisk protein som hører til F-boksen protein som en subunit av SCF E3 ubiquitin ligase protein kompleks (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005), I tillegg til TIR1, det er ytterligere tre F-boksen proteiner nemlig AUXIN UTTRYKK for at F-BOKSEN Proteiner (AFBs) som viser auxin-bindende aktivitet og formidle auxin signalering i Arabidopsis (Badescu og Napier, 2006). TIR1 reseptor kan samhandle med en gruppe av AUX/IAA (auxin/indole-3-eddiksyre) proteiner (Dharmasiri et al., 2003)., AUX/IAA proteiner er negative regulatorer av auxin signalering og det er 29 medlemmer av AUX/IAA kodet i Arabidopsis genom. AUX/IAA proteiner kan interagere med klasse av transcriptional regulatorer, auxin svar faktorer (ARF), til å formidle transcriptional svar til auxin. Under høy auxin nivå, AUX/IAA proteiner samhandle med TIR1 som coreceptor av auxin, og kan være ubiquitinated av SCFtir1 komplekse og dermed bli svekket gjennom ubiquitin–proteasome vei (Gray et al. I 2001; Lanza et al., 2012)., Ved ødeleggelse av AUX/IAA repressors, den auxin transcriptional regulatorer ARFs som inkluderer 23 medlemskap kan bli løst fra AUX/IAA undertrykkelse og dermed megle auxin svar ved aktivering eller undertrykkelse av mål gener (Guilfoyle og Hagen 2007). Forskjellige sett av F-boksen protein og AUX/IAA eller ARFs utlede kompleksiteten i løpet av auxin signal transduksjon (Goh et al., 2012; Guilfoyle, 2015; Salehin et al., 2015).,
koordinert handling av Aux/IAA transcriptional repressors og ARF transkripsjonfaktorer produserer komplekse gen-regulatoriske nettverk som også ble rapportert i Physcomitrella (Lavy et al., 2016). Nylig ble det funnet at CULLIN1 (CUL1) subunit av SCF i samspill med TIR1 og dermed regulerer TIR1 underlag stabilitet og auxin signalering (Wang et al., 2016). Samspillet mellom TIR1 og Aux/IAA er også påvirket av den romlige konformasjon av Aux/IAAs, kontrollert av en cyclophilin isomerase LRT2 i ris (Jing et al., 2015)., VARME SJOKK FAKTOR 90 (HSP90) og co-anstandsdame SGT1, henholdsvis, samhandler med TIR1 og dermed regulerer TIR1 stabilitet, noe som påvirker interaksjoner mellom TIR1 og Aux/IAA og auxin signalering (Wang et al., 2016).
Foruten TIR1 avhengige av kanonisk auxin-signalering vei, auxin har nylig blitt rapportert å lokke fram et variert utvalg av utviklingsmessige svar gjennom en ikke-kanoniske auxin-signalering mekanisme., I denne ikke-kanoniske auxin sensing prosessen, ARF3/ETTIN styrer genuttrykk gjennom interaksjoner med prosess-spesifikke transkripsjon faktorer, som svært beriker auxin-mediert anlegg utviklingsmessige mangfold (Simonini et al., 2016, 2017).
BR Signaliserer Sti
BRASSINOSTEROID ble først oppdaget i pollen for sin evne til å fremme celle strekning. Senere ble det funnet at BR spiller roller i et bredt spekter av plantevekst aspekter og kan svare på biotiske og abiotiske stressfaktorer., I dag BR signal transduksjon veien i stor grad var avklart med kombinasjoner av ulike metoder, inkludert molekylær genetikk, biokjemi, proteomics, og genomics, etc. Celle-overflaten kinase BRASSINOSTEROID INSENSITIVE1 (BRI1) ble identifisert som reseptoren av BR som kan binde seg til den ekstracellulære domenet av BRI1 og aktivere sin kinase aktivitet og dermed slå på et signal kaskade å regulere transkripsjon (Li og Chory, 1997; Wang et al. I 2001; Kinoshita et al., 2005; Kim og Wang, 2010; Clouse, 2011; Hothorn et al., 2011; Hun et al., 2011; Oh et al., 2012)., Ved oppfatningen av BR, BRI1 samhandler med co-reseptor BRI1-FORBUNDET KINASE 1 (BAK1) og dens homolog SOMATISKE EMBRYOGENESISRECEPTOR KINASEs (SERKs) for å danne en mer aktiv BR reseptor kompleks (Li et al., 2002; Nam og Li, 2002; Wang et al., 2005; Tang et al., 2008; Gou et al., 2012). Aktivert BRI1 phosphorylates to underlag av plasma membran-forankret reseptor-som cytoplasma kinases: BRASSINOSTEROID-SIGNALERING KINASES1 (BSK1) og KONSTITUTIVE DIFFERENSIAL GROWTH1 (- CDG1) (Tang et al., 2008; Kim et al.,, 2011), som i sin tur phosphorylates en PP1-type fosfatase heter BRI1-SUPPRESSOR1 (BSU1) for å aktivere BSU1, som fører til BSU1 dephosphorylation og inaktivering av GSK3-som kinase BRASSINOSTEROID INSENSITIVE2 (BIN2). Den kinase aktivitet av BIN2 er også hemmet av HISTONE DEACETYLASE HDA6, som samhandler og deacetylates på K189 av BIN2., Når BR nivåene er lave, BRI1 er hvilestrøm på grunn av sin negative regulator, BRI1 KINASE INHIBITOR 1 (BKI1) og protein fosfatase 2A (PP2A), mens BIN2 phosphorylate to BR homologe transkripsjonfaktorer, BRASSINAZOLE RESISTANT1 (BZR1) og BZR2 (også kalt BES1 for BRI1-EMS-SUPPRESSOR 1) (Han et al., 2002; Wang et al., 2002; Yin et al., 2002; Mora-Garcia et al., 2004; Kim et al., 2009, 2011; Kim og Wang, 2010). Når BR nivået er høyt, BIN2 er inaktivert, og BZR1 og BZR2 er dephosphorylated av PP2A, og flytte inn i kjernen til å endre uttrykket av tusenvis av BR respons-gener (Han et al.,, 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011).
Synergi Mellom BR og Auxin Signaliserer
Auxin og BR signal trasé spille ulike roller, men de viste også gjensidig forsterkende og avhengige av hverandre vekselsvirkningene i et bredt spekter av utviklingsmessige prosesser. For eksempel, både auxin og BR-signaler kan fremme celle ekspansjon og kan arbeide sammen for å fremme hypocotyls tøyelighet (Nemhauser et al., 2004)., Responsen fra en av de to veier for å fremme hypocotyl tøyelighet krever funksjonen av den andre og den gjensidige avhengigheten mellom BR og auxin trasé (Nemhauser et al., 2004). Auxin økt hypocotyl lengde i wild-type planter, men ikke i BR-ufølsom mutant bri1-116, og dette auxin-ufølsom fenotypen av bri1-116 var undertrykt av den dominerende gain-of-function mutant bzr1-1D, noe som indikerer BR eller aktiv BZR1 er nødvendig for auxin markedsføring av hypocotyl strekning., Det har blitt funnet at BR signaliserer konvergerer med SUPPRESSOR AV PHYTOCHROME B4-3 (SOB3) til å kontrollere celle tøyelighet og hypocotyl vekst gjennom regulering av auxin indusert SMÅ AUXIN OPP RNA19 (SAUR19) uttrykk (Favero et al., 2017). På den annen side, auxin regulert transkripsjonsfaktor LITE ORGAN STØRRELSE 1 (SMOS1) har nylig blitt funnet å kontrollere celle ekspansjon gjennom direkte interaksjon med SMOS2/DLT, medlem av GRAS familie av transcriptional co-regulatorer som spiller en positiv rolle i BR signalering i ris (Kim et al., 2009; Tong et al., 2012; Hirano et al., 2017)., Auxin i slekt mutanter som iaa3 og arf6/arf8 var mindre følsom for BR enn vill-type for hypocotyl tøyelighet, og avskaffet den overfølsomhet av bzr1-1D å auxin, noe som tyder på BR og BZR1 markedsføring av hypocotyl tøyelighet krever ARF6/8. Den genome-wide ChIP-Seq analyse avslørte at ARF6 aksjer et stort antall av genomisk mål (rundt 50%) med BZR1 og lys/temperatur-regulert transkripsjonsfaktor PIF4 av CHIP-Seq analyser (Oh et al., 2014)., BZR1 og PIF4 samhandle med ARF6 og aktivere felles mål gener ved å binde seg til felles mål gener fellesskap, under hypocotyls tøyelighet (Oh et al., 2014) og mange av disse overlappende mål gener som koder for cellevegg proteiner involvert i celle-utvidelsen.
Brassinosteroid og auxin også spille en viktig rolle i opprettholdelsen av rot apical meristem (RAM) (Durbak et al., 2012). RAM består av en liten gruppe sjelden dele celler kjent som hvilestrøm center (KK), omgitt av stamceller som kan gi opphav til ulike tann vev typer., Vedlikehold av roten stilk cellen befolkningen er regulert av WUSCHEL-RELATERTE HOMEOBOX 5 (WOX5) (Sarkar et al., 2007). WOX5 er begrenset til QC av auxin signalering og sikrer riktig uttrykk for PLT gener (Aida et al., 2004; Ding og Friml, 2010). Mutasjoner i BR reseptor genet BRASSINOSTEROID UFØLSOM 1 (BRI1) resultere i avvikende celle syklus progresjon i RAM og føre til en mindre RAMs (Gonzalez-Garcia et al., 2011; Hacham et al., 2011). Auxin er kjent for å stimulere biosyntese av BR (Chung et al.,, 2011), men aktiviteten av BR ikke påvirker uttrykket av PIN-gener (Hacham et al., 2011). Roten tips phenotypes av BR mutanter viser ikke den samme som den auxin mutanter (Gonzalez-Garcia et al., 2011), noe som indikerer at BR handle på RAM-uavhengig av auxin.
Brassinosteroid og auxin signaler er også synergi som kreves i radial mønster dannelsen av vaskulær bunter (Ibanes et al., 2009)., Ved kombinasjoner av matematisk modellering og biologiske eksperimenter, auxin maxima, som er etablert av asymmetrisk auxin polar transport, men ikke endringer på auxin nivåer er viktig for posisjonering i det vaskulære bunter. BR-signalet ble vist seg å fungere som et signal for å fremme antallet celler i den provascular ring som er i samsvar med auxin maxima. Dermed etablering av periodiske arrangement av vaskulær bunter i skyte er under koordinert handling av disse to anlegg hormoner (Ibanes et al., 2009)., Begge signalene er også involvert i anlegget rot utvikling og samspillet mellom BR og auxin er mediert av BREVIS RADIX (BRX) i løpet av denne prosessen. BRX er viktig for pris-begrense biosyntese av BR og BR eksogene programmet kan redde brx mutant feil. Videre, auxin-responsive genuttrykk er globalt svekket i brx mutant, og uttrykket av BRX er sterkt indusert av auxin og undertrykt av BR, noe som tyder på BR biosyntese og auxin signalering er koblet til via en feedback loop som involverer BRX under roten utvikling (Mouchel et al., 2006).,
Brassinosteroids og auxin også spille synergistisk roller under lateral rot utvikling. Baremqne i hovedsak fungere på den laterale rot primordia initiation mens auxin er nødvendige for både initiering og fremveksten stadier av lateral root-formasjonen (Casimiro et al. I 2001; Bhalerao et al., 2002; Benkova et al., 2003; Bao et al., 2004). I løpet av disse prosessene, Baremqne øke LRP initiering ved å fremme acropetal auxin transport i roten, men ikke ved å påvirke endogene IAA nivå (Bao et al., 2004)., Alle disse rapportene tyder på at krysstale mellom BR og auxin spiller en viktig rolle i reguleringen plante vekst og utvikling.
BR Regulerer Auxin Signaliserer
i Tillegg til den gjensidige avhengigheten og samarbeid av auxin og BR signaler anlegg under utvikling, BR kunne megle auxin signal vei på flere nivåer. BZR1 samhandler med ARF proteiner til direkte mål flere auxin signaliserer komponenter og gener som er involvert i auxin metabolisme, som transport og signalering, inkludert AUX/IAA, PINs, TIR1, og ARFs, etc. (Sun et al., 2010)., Det ble funnet at Aux/IAA proteiner er involvert i BR svar og iaa7/axr2-1 og iaa17/axr3-3 mutanter viste avvikende BR følsomhet og avvikende BR-indusert genuttrykk i et organ-avhengig måte (Nakamura et al., 2006). Eksogene brassinolide (BL) behandling kan påvirke uttrykket av auxin-responsive gener som IAA5, IAA19, IAA17, etc. og uttrykket ovenfor gener er nedregulert i BR biosynthetic mutant de-etiolated2 (det2), noe som indikerer at funksjonelle BR biosyntese er delvis kreves for auxin avhengige av genuttrykk (Nakamura et al.,, 2003; Kim et al., 2006). I tillegg, BR påvirker også auxin flyt ved å regulere uttrykk for auxin eksportører som PIN4 og PIN7 (Nakamura et al., 2004). I løpet av anlegget gravitropism svar, Baremqne kunne forbedre polar opphopning av auxin eksportør PIN2 i roten meristem sone, og dermed påvirke den omfordeling av auxin fra roten tips mot tøyelighet soner og resultat i forskjellen på IAA nivåer i både øvre og nedre sider av røtter for å indusere anlegg gravitropism., I løpet av denne prosessen, BR aktivert ROP2 spiller en viktig rolle i modulerende funksjonell lokalisering av PIN2-gjennom regulering av montering/demontering av F-actins (Li et al., 2005). Videre studier viste at redusert BL persepsjon og/eller konsentrasjon kan forårsake CYP79B2, genkode et enzym konvertering av tryptofan til indole-3-acetaldoxime og dermed påvirke fordelingen (Kim et al., 2007).
I tillegg ble det funnet at BR signal kunne regulere auxin signaliserer utgang av sin negative regulator GSK3 kinase BIN2., Den auxin svar faktor ARF2 ble identifisert som en BIN2 samspill protein i en gjær-to-hybrid tv og kinase analysen viste BIN2 kunne phosphorylate ARF2. Den phosphorylation av ARF2 resulterer i tap av sine DNA-bindende evne og undertrykkelse aktivitet i mål-gener (Vert et al., 2008). ARF2 er en BZR1 mål gener og dens uttrykk er redusert med BR behandling (Sun et al., 2010)., I tillegg, BIN2 kan phosphorylate ARF7 og ARF19 til å undertrykke sin interaksjon med AUX/IAAs og dermed forbedre transcriptional aktivitet på målet gener LATERAL ORGAN GRENSER-DOMAIN16 (LBD16) og LBD29 å regulere lateral rot organogenesis (Cho et al., 2014). Imidlertid, BR spiller en mindre rolle i denne prosessen og BIN2 er under kontroll av TRACHEARY ELEMENT DIFFERENSIERING HEMMENDE FAKTOR (TDIF)–TDIF RESEPTOR (TDR) modul (Cho et al., 2014). Sammen, BR kan regulere auxin reponses gjennom å påvirke ulike auxin signaliserer komponenter.,
Auxin Regulerer BR Signaliserer
På den annen side, auxin kan også regulere BR signal vei i visse aspekter. Uttrykket av DWARF4, en avgjørende hydroksylase for BR biosyntese å kontrollere endogene BR nivå, er auxin avhengige. Auxin behandling kunne merkbart stimulere uttrykk for DWARF4 og auxin kan hemme bindingen av BZR1 arrangøren av DWARF4. Induksjon av DWARF4 av auxin krever auxin signaliserer veien, men ikke BR signaliserer vei (Chung et al., 2011; Yoshimitsu et al., 2011)., CPD catalyzing C-3 oksidasjon av BR ble aktivert av BRX, antatte en transkripsjonsfaktor som handler nedstrøms av auxin signalering (Mouchel et al., 2006). Videre studier i ris indikerer at eksogene auxin kan forbedre transkripsjon uttrykk nivåer av BR reseptor genet OsBRI1, noe som tyder på at auxin forbedrer BR signalering gjennom regulering av BR-reseptorer (Sakamoto et al., 2013). Videre formidler av OsBRI1 har en oppstrøms auxin-respons-element (AuxRE) motiv som er målrettet ved ARF transkripsjon faktorer., Videre mutant studier indikerer at ved mutasjon av AuxRE, induksjon av uttrykk av OsBRI1 av auxin er avskaffet og også uttrykk for OsBRI1 er nede regulert i arf mutant (Sakamoto et al., 2013). Det har blitt rapportert at OsARF19 binder seg til arrangøren av OsBRI1 og positivt regulerer sitt uttrykk som deretter aktiverer BR signalering (Zhang et al., 2015). BES1 kan binde seg til arrangøren av SMÅ AUXIN-UP-RNA-15 (SAUR15) og formidle BR tidlig respons genet i Arabidopsis, og denne bindingen kan være forsterket av auxin behandling (Walcher og Nemhauser, 2012)., Tatt sammen, auxin kan også påvirke BR svar og BR regulert plante vekst og utvikling.
Avsluttende Bemerkninger og Fremtidig Perspektiv
i Løpet av de siste nesten fire tiår, studier på auxin-BR vei vekselsvirkningene har tiltrukket flere og flere forskere’ interesse. Apparatet av fysiologiske, molekylære genetiske og biokjemiske verktøy har i stor grad utdypet vår forståelse av dette problemet., Basert på tidligere studier, BR og auxin er involvert sammen i flere anlegg utviklingsmessige prosesser, inkludert: hypocotyl tøyelighet, vaskulær bunter utvikling, rot utvikling og tropisms, etc. Den gjensidige avhengigheten og samarbeid av auxin og BR er komplisert og involverer mange prosesser på molekylært nivå, ved å dele samme mål gener som regulerer hverandre på flere nivåer (Figur 1).
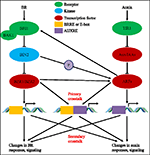
FIGUR 1. Modell av auxin-brassinosteroid (BR) krysstale., I Arabidopsis, oppfatninger av BR og auxin signal er gjenkjent av BRI1 og TIR1 reseptorer, henholdsvis. BR binder seg til den ekstracellulære domenet av BRI1 og fremme den kommuniserer med co-reseptor BAK1 å danne en mer aktiv BR reseptor kompleks, som i sin tur føre til dephosphorylation og inaktivering av BIN2. Den inaktivering av BIN2 føre til dephosphorylation av to BR homologe transkripsjonfaktorer BZR1 og BZR2, som flytter inn i kjernen for å aktivere transkripsjon av gener som inneholder BRRE eller E-boks i sin promoter-regionen., BIN2 også kan phosphorylate ARF7 og ARF19 til å undertrykke sin interaksjon med AUX/IAAs og dermed forbedre transcriptional aktivitet på målet gener. TIR1 mottak av auxin signalering og samhandle med AUX/IAA proteiner som co-reseptoren av auxin. AUX/IAA så er forringet gjennom ubiquitin–proteasome vei, og auxin transcriptional regulatorer auxin svar faktorer (ARFs) er utgitt fra AUX/IAA undertrykkelse og aktivere transkripsjon av gener med auxin responsive elementer (AUXRE) i deres regulerende regionen., Noen ARFs kan også binder seg til arrangøren av BRI1 og positivt regulerer sitt uttrykk som deretter aktiverer BR signalering. Primære krysstale skjer ved aktivering av gener som inneholder både BRRE/E-boksen og AUXRE i deres promoter-regionen, slik at både signaliserer veier til direkte regulerer transkripsjon. Sekundær krysstale skjer gjennom uttrykk av gener som enten er auxin eller BR responsiv, men aktiviteter som kontrollerer uttrykket av gener som regulerer respons og signalering av andre hormoner.,
Phosphorylation regulering spiller en avgjørende rolle i BR signaliserer veien, spesielt i løpet av den oppfatning prosessen, BR oppfattes gjennom BRI1 kinase reseptor og BAK1 kinase co-reseptorer, og til slutt kontroller BR regulert genuttrykk gjennom å påvirke nedstrøms transkripsjon faktorer som BES1/BZR1 aktiviteter (Han et al., 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011). Imidlertid, ubiquitination regulering synes avgjørende for auxin signalering., Når auxin binder seg til TIR1 reseptor, som fungerer som et ubiquitin E3-ligase, den aktiverte TIR1 E3-ligase ubiquitinates AUX/IAA proteiner, fører til nedbrytning av disse repressors og de-undertrykker ARF transkripsjon faktorer, og til slutt fører til auxin regulert genuttrykk mønster endringer og vekst svar (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005). Siden det har blitt funnet at BIN2 kinase, som er godt kjent funksjon i BR signalering, kan phosphorylate og forbedre aktivitetene ARFs som ARF2 og ARF7 (Vert et al., 2008; Cho et al.,, 2014), vil det være interessant å teste om kinases som BIN2, som er involvert i BR signalering, kan også samhandle med andre auxin signaliserer komponenter som TIR1 reseptor eller AUX/IAA repressors og innflytelse TIR1 E3-ligase aktivitet eller AUX/IAA protein stabilities. På den annen side, er den rollen som ubiquitination i BR signaliserer også behov for å bli behandlet, spesielt hvis TIR1 E3-ligase kunne samhandle direkte med BR signaliserer komponenter og regulere deres protein stabilities.,
I tillegg, ved hjelp av auxin svar DR5 og andre auxin journalister, har det vært observert at auxin regulerer plantevekst og utvikling i et vev-eller mobiltelefonnett-avhengig måte. De ulike transcriptional utganger avhengig av mobil-og miljømessige konteksten (Clark et al., 2014; Etchells et al., 2016; Lavy et al., 2016)., Selv om spatiotemporal BR signaliserer har vist seg å kontrollere rotvekst gjennom den antagonistiske handling med auxin (Chaiwanon og Wang, 2015), det er fortsatt ukjent om vev eller mobil BR signalering, som kan bli visualisert ved pBZR1:BZR1-YFP, er også viktig for å styre andre prosesser i tillegg til roten utvikling. Videre generering av en detaljert vev eller kjeller kart av auxin og BR-distribusjoner som er mulig i dag ved hjelp av fluorescens-aktivert celle-sortering eller laser microdissection i kombinasjon med høy oppløsning genuttrykk analyse., Dette vil til slutt fører til adressen hvis auxin crosstalks med BR i en vev eller et mobilnettverk måte.
Forfatter Bidrag
Alle forfatterne var involvert i skrivingen av denne anmeldelsen manuskriptet. For mer informasjon om hva som utgjør forfatterskap, vennligst se vår forfatter retningslinjer.,
Midler
Dette arbeidet er støttet med midler fra National Natural Science Foundation of China (Prosjekter 31500229 og 31770305), Shandong-Provinsen Natural Science Foundation of Major Grunnleggende Research Program (2017C03), Shandong Provinsielle Midler for Fremragende Unge Forskere (2014JQ201408).
interessekonflikt Uttalelse
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller finansielle forhold som kan oppfattes som en potensiell interessekonflikt.
Clouse, S. D. (2011)., Brassinosteroid signal transduksjon: fra reseptor kinase aktivering å transcriptional nettverk regulere anlegget utvikling. Plante Celle 23, 1219-1230. doi: 10.1105/tpc.111.084475
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar
Estelle, M. (2011). Auxin Signalering: Fra Syntese Systemer Biologi; Et Emne Samling fra Cold Spring Harbor Perspektiver i Biologi. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Trykk.
Google Scholar
Guilfoyle, T. J. (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar