introductie
In tegenstelling tot dieren die kunnen bewegen om de ongunstige omgeving te vermijden, vertonen de sessiele planten een sterk ontwikkelde aanpassing aan de ingewikkelde omgevingsomstandigheden. Om dit diepgaande aanpassingsvermogen te bereiken, zijn communicatie tussen cellen noodzakelijk. Cel aan celcommunicatie in installaties impliceert robuuste intracellular signalerende verwerking en ingewikkelde intercellulaire signalerende netwerken., Tot nu toe zijn er ten minste negen signalerende stoffen, genaamd plantenhormonen, waaronder auxine, brassinosteroïde (BR), cytokinine, gibberellinen (GA), ethyleen, jasmoninezuur (JA), strigolacton (SL), abscisinezuur (ABA), en salicylzuur (SA) ontdekt (Druege et al., 2016; Verma et al., 2016). De genetische en fysiologische studies hebben de kritieke rollen en de functionele mechanismen van deze bovengenoemde hormonen in de groei en ontwikkeling van planten onthuld (Gray, 2004)., Op basis van de voorgaande studies functioneren auxine, BR, GA, SL en cytokinine voornamelijk tijdens de normale groei en ontwikkeling van planten, terwijl ABA, ethyleen, JA en SA een belangrijke rol spelen in de groeireactie van planten op verschillende biotische en abiotische stress (Pieterse et al., 2009; Santner et al., 2009; Denance et al., 2013). En ook sommige van deze hormonen hebben dubbele rollen, bijvoorbeeld, ABA speelt ook belangrijke rollen in zaadontwikkeling en rustperiode (Seo en Koshiba, 2002)., Hoewel elk hormoon overheersende rollen in bepaalde aspecten speelt, hebben vele hormonen overlappende activiteiten en de interactie van verschillende hormonen controle vele ontwikkelingsaspecten en groei in reactie op endogene ontwikkelings en exogene signalen.
auxine en BR zijn twee belangrijke klassen groeibevorderende hormonen., BR, Een groep plantspecifieke steroïde hormonen die kunnen interageren met andere fytohormonen zoals auxine, cytokinine, ethyleen, GA, JA en SA en een breed scala aan plantengroei-en ontwikkelingsprocessen reguleren, waaronder zaadkieming, celverlenging, vasculaire differentiatie, stomataformatie en beweging, bloei en mannelijke vruchtbaarheid (Saini et al., 2015). Interessant is dat elk van deze processen ook wordt gecontroleerd door auxine, wat suggereert dat deze twee hormonen samenwerken om de ontwikkeling van planten te controleren., In dit overzicht zullen we de signaaltransductie van auxine en BR schetsen op basis van de recente vooruitgang en de kruisverwijzing tussen auxine en BR gemedieerde plantengroei en-ontwikkeling bekijken.
Auxine signaalweg
Auxine werd voor het eerst herkend als planthormoon vanwege zijn rol in plant tropisme op zwaartekracht-of lichtstimuli., Later werd auxine chemisch geïdentificeerd als indool-3-azijnzuur en bleek het een essentiële rol te spelen in een overvloed aan plantenontwikkelings-en fysiologische processen, waaronder embryogenese, organogenese, vasculaire differentiatie, wortel-en scheutontwikkeling, tropische groei en fruitontwikkeling (Estelle, 2011).
met behulp van genetische analyse in Arabidopsis is het moleculaire mechanisme dat ten grondslag ligt aan de auxine signaaltransductie goed onderzocht. TRANSPORT INHIBITOR RESPONSE1 (TIR1) was de eerste geïdentificeerde nucleaire receptor van auxine (Ruegger et al., 1998; Dharmasiri et al., 2005)., TIR1 codeert een kernproteïne die tot het F-box-eiwit behoort als subeenheid van SCF E3 ubiquitin ligase – eiwitcomplex (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005), naast TIR1, zijn er drie extra F-box proteã nen namelijk auxine signalerende F BOX Proteã nen (AFBs) die auxine-bindende activiteit tonen en auxine signalerende in Arabidopsis medieren (Badescu en Napier, 2006). TIR1-receptor kan interageren met een groep Aux/IAZ (auxine / indool-3-azijnzuur) eiwitten (Dharmasiri et al., 2003)., AUX / IAZ-eiwitten zijn negatieve regulatoren van auxine-signalering en er zijn 29 leden van AUX/IAZ gecodeerd in het Arabidopsis-genoom. AUX / IAZ-eiwitten kunnen interageren met de klasse van transcriptionele regulatoren, auxineresponsfactoren (ARF), om transcriptionele responsen op auxine te mediëren. Onder een hoog auxinegehalte interageren AUX/IAA-eiwitten met TIR1 als coreceptor van auxine, en kunnen ze worden ubiquitinated door het scftir1-complex en dus worden afgebroken via de ubiquitine-proteasoomroute (Gray et al., 2001; Lanza et al., 2012)., Na de vernietiging van AUX/IAA-onderdrukkers kunnen de auxine transcriptionele regulatoren ARF ‘ s, die 23 lidmaatschappen omvatten, worden bevrijd van Aux/IAA-onderdrukking en zo de auxine-reacties bemiddelen door activering of onderdrukking van doelgenen (Guilfoyle en Hagen, 2007). De verschillende sets van F-box proteïne en AUX / IAA of ARFs concluderen de complexiteit tijdens auxine signaaltransductie (Goh et al., 2012; Guilfoyle, 2015; Salehin et al., 2015).,
de gecoördineerde werking van Aux / IAZ transcriptionele onderdrukkers en ARF transcriptiefactoren produceert complexe genregulerende netwerken die ook werden gerapporteerd in Physcomitrella (Lavy et al., 2016). Onlangs, werd gevonden dat CULLIN1 (CUL1) subeenheid van het SCF met TIR1 in wisselwerking staat en zo tir1 substraten stabiliteit en auxine signaleren regelt (Wang et al., 2016). De interactie tussen TIR1 en Aux / IAA wordt ook beïnvloed door de ruimtelijke bouw van Aux/ IAAs, gecontroleerd door een cyclophiline-isomerase LRT2 in rijst (Jing et al., 2015)., Hitteschok FACTOR 90 (HSP90) en de co-chaperone SGT1, respectievelijk, interageert met TIR1 en reguleert zo TIR1 stabiliteit, die de interacties tussen TIR1 en Aux/IAA En auxine signalering beïnvloedt (Wang et al., 2016).
naast de TIR1-afhankelijke canonieke auxine-signaalweg, is onlangs gemeld dat auxine een breed scala aan ontwikkelingsreacties uitlokt via een niet-canoniek auxine-signaalmechanisme., In dit niet-canonieke auxine sensing proces controleert ARF3/ETTIN genexpressie door interacties met process-specific transcriptiefactoren, die auxine-gemedieerde plantenontwikkelingsdiversiteit sterk verrijken (Simonini et al., 2016, 2017).
br signaalweg
BRASSINOSTEROÏDE werd voor het eerst ontdekt in pollen vanwege zijn vermogen om celelongatie te bevorderen. Later bleek dat BR rollen speelt in een breed scala van plantengroei aspecten en kan reageren op biotische en abiotische spanningen., Tegenwoordig werd de signaaltransductieweg van BR grotendeels verduidelijkt door combinaties van verschillende methodes, met inbegrip van moleculaire genetica, biochemie, proteomica, en genomica, enz. De cel-oppervlakte kinase BRASSINOSTEROID ONGEVOELIGHEID1 (BRI1) werd geà dentificeerd als receptor van BR die aan het extracellulaire domein van BRI1 kan binden en zijn kinaseactiviteit activeren en zo een signaalcascade inschakelen om transcriptie te regelen (Li en Chory, 1997; Wang et al., 2001; Kinoshita et al., 2005; Kim and Wang, 2010; Clouse, 2011; Hothorn et al., 2011; She et al., 2011; Oh et al., 2012)., Na waarneming van BR interageert BRI1 met Co-receptor BRI1-geassocieerd KINASE 1 (BAK1) en zijn homolog somatische EMBRYOGENESISRECEPTOR KINASEs (SERKs) om een actiever br-receptorcomplex te vormen (Li et al., 2002; Nam and Li, 2002; Wang et al., 2005; Tang et al., 2008; Gou et al., 2012). Geactiveerde BRI1 fosforylaten twee substraten van plasmamembraan-verankerde receptor-achtige cytoplasmatische kinasen: BRASSINOSTEROID-signalerende KINASES1 (BSK1) en CONSTITUTIVE DIFFERENTIAL GROWTH1 (CDG1) (Tang et al., 2008; Kim et al.,, 2011), die op zijn beurt phosphorylates een PP1-type fosfatase genoemd BRI1-ONDERDRUKKER1 (BSU1) om bsu1 te activeren, leidend tot bsu1 dephosphorylation en inactivatie GSK3-als kinase BRASSINOSTEROID ONGEVOELIGHEID2 (BIN2). De kinaseactiviteit van BIN2 wordt ook geremd door HISTONE DEACETYLASE HDA6, die in wisselwerking staat en deacetylaten bij K189 van BIN2., Wanneer BR niveaus zijn laag, BRI1 is bevroren vanwege de negatieve regulator, BRI1 KINASE INHIBITOR 1 (BKI1) en proteïne fosfatase 2A (PP2A), terwijl BIN2 phosphorylate twee BR homologe transcriptie factoren, BRASSINAZOLE RESISTANT1 (BZR1) en BZR2 (ook genoemd BES1 voor BRI1-EMS-SUPPRESSOR 1) (He et al., 2002; Wang et al., 2002; Yin et al., 2002; Mora-Garcia et al., 2004; Kim et al., 2009, 2011; Kim en Wang, 2010). Wanneer BR-niveaus hoog zijn, wordt BIN2 geïnactiveerd, en BZR1 en BZR2 worden gedefosforyleerd door PP2A, en bewegen zich in kern om de uitdrukking van duizenden br-reactiegenen te veranderen (he et al.,, 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011).
de synergie tussen br en Auxine signaalwegen
auxine en BR signaalwegen spelen verschillende rollen, maar ze toonden ook synergetische en onderling afhankelijke interacties in een breed scala van ontwikkelingsprocessen. Zowel auxine-als BR-signalen kunnen bijvoorbeeld celuitbreiding bevorderen en synergistisch interageren om hypocotylen-verlenging te bevorderen (Nemhauser et al., 2004)., De respons van een van de twee routes in het bevorderen van hypocotyl elongatie vereist de functie van de andere en de onderlinge afhankelijkheid tussen br en auxine routes (Nemhauser et al., 2004). Auxine verhoogde de hypocotyllengte in wild-type planten maar niet in de BR-ongevoelige mutant bri1-116, en dit auxine-ongevoelige fenotype van bri1-116 werd onderdrukt door de dominante gain-of-function mutant bzr1-1D, wat aangeeft dat BR of actieve BZR1 nodig is voor auxine promotie van hypocotyl elongatie., Het is gebleken dat br-signalering convergeert met SUPPRESSOR van Fytochroom B4-3 (SOB3) om celelongatie en hypocotylgroei te controleren door de regulering van auxine geïnduceerde kleine auxine up RNA19 (SAUR19) expressie (Favero et al., 2017). Aan de andere kant, de auxine gereguleerde transcriptiefactor klein orgaan Grootte 1 (SMOS1) is onlangs gevonden om celuitbreiding te controleren door de directe interactie met SMOS2/DLT, een lid van de GRAS familie van transcriptionele co-regulators die een positieve rol speelt in BR signalering in rijst (Kim et al., 2009; Tong et al., 2012; Hirano et al., 2017)., Auxine gerelateerde mutanten zoals iaa3 en arf6/arf8 waren minder gevoelig voor BR dan wild-type voor hypocotyl elongatie, en schaften de overgevoeligheid van bzr1-1D voor auxine af, wat suggereert dat de bevordering van BR en BZR1 van hypocotyl elongatie arf6/8 vereist. De genoom-brede Spaander-Seq analyse openbaarde dat ARF6 een groot aantal genomic doelstellingen (rond 50%) met BZR1 en het licht/temperatuur-geregelde transcriptiefactor PIF4 door spaander-Seq analyses deelt (Oh et al., 2014)., BZR1 en PIF4 interageren met ARF6 en activeren gedeelde doelgenen door samen te binden aan gedeelde doelgenen tijdens hypocotylen verlenging (Oh et al., 2014) en veel van deze overlappende doelgenen coderen celwandproteã nen betrokken bij celuitbreiding.Brassinosteroïde en auxine spelen ook een belangrijke rol in het behoud van apicale meristem (RAM) (Durbak et al., 2012). De RAM bestaat uit een kleine groep van zelden delende cellen bekend als het rustige centrum (QC), omgeven door stamcellen die aanleiding geven tot de verschillende Toot weefseltypes., Het onderhoud van de stamcelpopulatie van de wortel wordt geregeld door WUSCHEL-gerelateerde HOMEOBOX 5 (WOX5) (Sarkar et al., 2007). WOX5 is beperkt tot de QC door auxine signalering en vergemakkelijkt de juiste expressie van de PLT genen (Aida et al., 2004; Ding and Friml, 2010). Mutaties in het BR-receptorgen BRASSINOSTEROÏDE ongevoelig 1 (BRI1) resulteren in afwijkende progressie van de celcyclus in de RAM en veroorzaken een kleinere Ram (Gonzalez-Garcia et al., 2011; Hacham et al., 2011). Van auxine is bekend dat het de biosynthese van BR stimuleert (Chung et al.,, 2011), maar de activiteit van BR heeft geen invloed op de expressie van PIN genen (Hacham et al., 2011). De fenotypen van de worteltip van BR-mutanten vertonen niet hetzelfde als de auxine-mutanten (Gonzalez-Garcia et al., 2011), waaruit blijkt dat BR onafhankelijk van auxine op het RAM werkt.
Brassinosteroïde – en auxinesignalen zijn ook synergetisch nodig in de radiale patroonvorming van vasculaire bundels (Ibanes et al., 2009)., Door de combinaties van wiskundige modellering en biologische experimenten is auxine maxima, vastgesteld door asymmetrisch auxine polair transport, maar geen veranderingen op auxine niveaus belangrijk voor de positionering van de vasculaire bundels. Het BR-signaal bleek te dienen als een bevorderend signaal voor het aantal cellen in de provasculaire ring die consistent zijn met auxine maxima. Aldus is de vaststelling van periodieke rangschikking van vasculaire bundels in de scheut onder de gecoördineerde actie van deze twee plantaardige hormonen (Ibanes et al., 2009)., Beide signalen zijn ook betrokken bij de ontwikkeling van plantenwortels en de interactie van BR en auxine wordt gemedieerd door BREVIS RADIX (BRX) tijdens dit proces. BRX is belangrijk voor de snelheidsbeperkende biosynthese van BR en Br exogene toepassing kan brx mutantdefecten redden. Bovendien is de auxine-responsieve genexpressie wereldwijd verminderd in brx mutant, en de expressie van BRX wordt sterk geïnduceerd door auxine en onderdrukt door BR, wat impliceert dat de biosynthese van BR en de auxine-signalering verbonden zijn door een terugkoppelingslus waarbij BRX betrokken is tijdens de wortelontwikkeling (Mouchel et al., 2006).,
Brassinosteroïden en auxine spelen ook een synergetische rol tijdens de laterale wortelontwikkeling. BRs functioneren voornamelijk bij de laterale wortel primordia initiatie terwijl auxine nodig is voor zowel initiatie als uitkomststadia van laterale wortelvorming (Casimiro et al., 2001; Bhalerao et al., 2002; Benkova et al., 2003; Bao et al., 2004). Tijdens deze processen verhoogt BRs de LRP-initiatie door acropetaal auxinetransport in de wortel te bevorderen, maar niet door het endogene IAZ-niveau te beïnvloeden (Bao et al., 2004)., Al deze rapporten suggereren dat de kruisverwijzing tussen BR en auxine een belangrijke rol speelt in de regulerende plantengroei en-ontwikkeling.
BR reguleert Auxinesignaal
naast de onderlinge afhankelijkheid en samenwerking van auxine-en BR-signalen tijdens de ontwikkeling van de plant, kan BR auxinesignaalweg op meerdere niveaus bemiddelen. BZR1 staat in wisselwerking met ARF-proteã nen om veelvoudige auxine signalerende componenten en genen direct te richten betrokken bij auxinmetabolisme zoals vervoer en signalerend, met inbegrip van AUX/IAA, spelden, TIR1, en ARFs, enz. (Sun et al., 2010)., Er werd vastgesteld dat Aux / IAA-eiwitten betrokken zijn bij BR-responsen en dat iaa7/axr2-1 en iaa17 / axr3-3-mutanten afwijkende br-gevoeligheid en afwijkende BR-geïnduceerde genexpressie vertoonden op een orgaanafhankelijke manier (Nakamura et al., 2006). Exogene brassinolide (BL) behandeling kan de expressie van auxine-responsieve genen zoals iaa5, IAA19, IAA17, enz.induceren., en de expressie van de bovengenoemde genen is down-gereguleerd in de BR biosynthetische mutant de-etioated2 (det2), wat aangeeft dat functionele br biosynthese gedeeltelijk nodig is voor auxine-afhankelijke genexpressie (Nakamura et al.,, 2003; Kim et al., 2006). Bovendien beïnvloedt BR ook de auxinestroom door de expressie van auxine-exporteurs zoals PIN4 en PIN7 te reguleren (Nakamura et al., 2004). Tijdens planten-gravitropismereacties kan BRs de polaire accumulatie van de auxine-exporteur PIN2 in de wortelmeristemzone versterken en zo de herverdeling van auxine van de worteltip naar de elongatiezones beïnvloeden en resulteren in het verschil in IAZ-niveaus in zowel de boven-als de onderzijde van de wortels om plant-gravitropisme te induceren., Tijdens dit proces, speelt br geactiveerde ROP2 een belangrijke rol in het moduleren van de functionele lokalisatie van PIN2 door de verordening van de assemblage / hermontage van F-actins (Li et al., 2005). Verdere studies toonden aan dat verminderde BL perceptie en / of concentratie CYP79B2 kon induceren, het gen dat codeert voor een enzym dat tryptofaan omzet in indool-3-acetaldoxim en zo de distributie beïnvloedt (Kim et al., 2007).
bovendien werd vastgesteld dat het BR-signaal auxine signaaluitgang kon regelen door de negatieve regulator gsk3 kinase BIN2., De auxine-responsfactor ARF2 werd geïdentificeerd als een BIN2-interagerend eiwit in een gist tweehybride screen en kinase assay toonde BIN2 kon fosforylaat ARF2. De fosforylering van ARF2 resulteert in het verlies van zijn DNA-bindingsvermogen en onderdrukkingsactiviteit van de doelgenen (Vert et al., 2008). ARF2 is een bzr1-doelgenen en de expressie ervan wordt verminderd door BR-behandeling (Sun et al., 2010)., Bovendien, BIN2 kan phosphorylate ARF7 en ARF19 om hun interactie met AUX/IAAs te onderdrukken en daardoor de transcriptional activiteit op hun doelgenen laterale ORGAANGRENZEN verbeteren-DOMAIN16 (LBD16) en LBD29 om laterale wortelorganogenese te regelen (Cho et al., 2014). Echter, BR speelt een kleine rol tijdens dit proces en BIN2 is onder de controle van de TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR (TDIF)–TDIF RECEPTOR (TDR) module (Cho et al., 2014). Samen kan BR auxine reponsen reguleren door verschillende auxine signaalcomponenten te beïnvloeden.,
Auxine reguleert BR-signalering
anderzijds kan auxine ook de BR-signaalroute in bepaalde aspecten reguleren. De expressie van DWARF4, een cruciale hydroxylase voor de biosynthese van BR om het endogene BR-niveau te controleren, is afhankelijk van auxine. Behandeling met auxine kan de expressie van DWERG4 merkbaar stimuleren en auxine kan de binding van BZR1 aan de promotor van DWERG4 remmen. De inductie van DWARF4 door auxine vereist auxine signaalweg maar niet BR signaalweg (Chung et al., 2011; Yoshimitsu et al., 2011)., CPD katalyserende C-3 oxidatie van BR werd geactiveerd door BRX, een vermeende transcriptiefactor die stroomafwaarts werkt van auxine signalering (Mouchel et al., 2006). Verdere studie in rijst geeft aan dat exogeen auxine de transcriptie expressieniveaus van BR receptor gen OsBRI1 kan verhogen, wat suggereert dat auxine br-signalering versterkt door de regulatie van BR-receptoren (Sakamoto et al., 2013). Bovendien bezit de promotor van OsBRI1 een upstream auxine-response element (AuxRE) motief dat gericht is op ARF transcriptiefactoren., Bovendien geven mutantstudies aan dat bij mutatie van AuxRE de inductie van expressie van OsBRI1 door auxine wordt afgeschaft en dat ook de expressie van OsBRI1 down gereguleerd wordt in arf mutant (Sakamoto et al., 2013). Er is gemeld dat OsARF19 bindt aan de promotor van OsBRI1 en positief zijn uitdrukking reguleert die dan de BR-signalering activeert (Zhang et al., 2015). BES1 kan binden aan de promotor van klein auxine-up RNA 15 (SAUR15) en Br vroege respons gen in Arabidopsis bemiddelen, en deze binding kan worden versterkt door auxine behandeling (Walcher and Nemhauser, 2012)., Samen kan auxine ook de BR-respons en de BR-gereguleerde plantengroei en-ontwikkeling beïnvloeden.
slotopmerkingen en toekomstperspectief
in de afgelopen vier decennia hebben studies naar auxine-BR-pathway-interacties steeds meer belangstelling van onderzoekers gewekt. Het gebruik van fysiologische, moleculaire, genetische en biochemische hulpmiddelen heeft ons begrip van deze kwestie aanzienlijk verdiept., Op basis van de voorgaande studies zijn BR en auxine synergistisch betrokken bij meerdere plantontwikkelingsprocessen, waaronder: hypocotyl elongatie, vasculaire bundels ontwikkeling, wortelontwikkeling en tropismen, enz. De onderlinge afhankelijkheid en samenwerking van auxine en BR zijn ingewikkeld en omvatten talrijke processen op moleculair niveau, door het delen van dezelfde doelgenen, die elkaar wederzijds op meerdere niveaus reguleren (figuur 1).
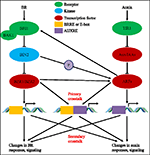
figuur 1. Model van auxin-brassinosteroid (BR) kruisverwijzing., In Arabidopsis worden de waarnemingen van BR en auxine signaal herkend door respectievelijk BRI1 en TIR1 receptoren. BR bindt aan het extracellulaire domein van BRI1 en bevordert het interageert met co-receptor BAK1 om een actiever br receptorcomplex te vormen, dat op zijn beurt tot de dephosphorylation en inactivatie van BIN2 leidt. De inactivatie van BIN2 leidt tot de dephosphorylatie van twee br homologe transcriptiefactoren BZR1 en BZR2, die in kern bewegen om transcriptie van genen te activeren die BRRE of E-box in hun promotorgebied bevatten., BIN2 kan ook phosphorylate ARF7 en ARF19 om hun interactie met AUX/IAAs te onderdrukken en daardoor de transcriptional activiteit op hun doelgenen verbeteren. TIR1 ontvangt de auxine-signalering en interageert met AUX/IAZ-eiwitten als co-receptor van auxine. De AUX/IAZ wordt vervolgens afgebroken via de ubiquitine-proteasoomroute, en de auxine transcriptionele regulatoren auxine responsfactoren (ARFs) worden vrijgegeven van Aux/IAZ onderdrukking en activeren transcriptie van genen met auxine responsieve elementen (AUXRE) in hun regulerende regio., Sommige ARFs kan ook aan de promotor van BRI1 binden en positief zijn uitdrukking regelen die dan br Het signaleren activeert. Primaire kruisverwijzing vindt plaats door activering van genen die zowel BRRE/E-box als AUXRE in hun promotorgebied bevatten, waardoor beide signaalwegen de transcriptie direct kunnen regelen. Secundaire kruisverwijzing komt voor door expressie van genen die auxine of BR reageren, maar waarvan de activiteiten de expressie van genen controleren die de respons en signalering van andere hormonen reguleren.,
Fosforyleringsregulatie speelt een cruciale rol in de BR-signaalroute, vooral tijdens het waarnemingsproces, wordt BR waargenomen via BRI1-kinasereceptor en BAK1-kinase-co-receptoren, en controleert uiteindelijk BR gereguleerde genexpressie door downstream transcriptiefactoren zoals BES1/BZR1-activiteiten te beïnvloeden (He et al., 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011). Ubiquitinatieregulering lijkt echter essentieel voor auxine-signalering., Zodra auxine zich bindt aan de TIR1-receptor, die als ubiquitine E3-ligase fungeert, leidt de geactiveerde TIR1 E3-ligase ubiquitineert AUX/IAA-eiwitten, tot de degradatie van deze onderdrukkers en de-onderdrukt ARF transcriptiefactoren, en veroorzaakt uiteindelijk auxine gereguleerde genexpressiepatronen en groeireacties (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005). Aangezien men heeft gevonden dat BIN2 kinase, dat goed gekend het functioneren in BR signaleren is, kon phosphorylate en de activiteiten van ARFs zoals ARF2 en ARF7 (Vert et al., 2008; Cho et al.,, 2014), zal het interessant zijn om te testen of kinases zoals BIN2, die betrokken zijn bij het signaleren van BR, ook met andere auxine signalerende componenten zoals TIR1 receptor of AUX/IAA-onderdrukkers kunnen interageren, en TIR1 E3-ligase activiteit of AUX/IAA eiwitstabiliteit kunnen beïnvloeden. Anderzijds, moet de rol van ubiquitination in BR het signaleren ook worden aangepakt, vooral als TIR1 E3-ligase direct met BR signalerende componenten kon in wisselwerking staan en hun eiwitstabiliteit regelen.,
daarnaast is, met behulp van auxine respons DR5 en andere auxine reporters, waargenomen dat auxine de groei en ontwikkeling van planten reguleert op een weefsel of Cellulaire afhankelijke manier. De diverse transcriptionele outputs afhankelijk van de cellulaire en omgevingscontext (Clark et al., 2014; Etchells et al., 2016; Lavy et al., 2016)., Hoewel is aangetoond dat de spatiotemporale br-signalering wortelgroei door de antagonistische werking met auxine (Chaiwanon en Wang, 2015) kan controleren, is het nog onbekend of het weefsel of de cellulaire br-signalering, die kan worden gevisualiseerd door pBZR1:BZR1-YFP, ook belangrijk is om andere processen naast wortelontwikkeling te controleren. Bovendien is het genereren van een gedetailleerde weefsel-of kelderkaart van auxine-en BR-distributies momenteel mogelijk met behulp van fluorescentie-geactiveerde celsortering of laser microdissectie in combinatie met een hoge-resolutie genexpressieanalyse., Dit zal uiteindelijk leiden tot address als de auxine kruist met BR op een weefsel of Cellulaire manier.
Auteursbijdragen
alle auteurs waren betrokken bij het schrijven van dit recensiemanuscript. Voor meer informatie over wat auteurschap is, verwijzen wij u naar onze auteursrichtlijnen.,
financiering
Dit werk wordt ondersteund door subsidies van de National Natural Science Foundation of China (projecten 31500229 en 31770305), Shandong Province Natural Science Foundation of Major Basic Research Program (2017C03), Shandong Provincial Funds for Distinguished Young Scholars (2014JQ201408).
belangenconflict verklaring
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
Clouse, S. D. (2011)., Brassinosteroid signaaltransductie: van receptor kinase activatie naar transcriptionele netwerken die de ontwikkeling van planten reguleren. Plant Cell 23, 1219-1230. doi: 10.1105 / tpc.111.084475
PubMed Abstract / CrossRef Full Text / Google Scholar
Estelle, M. (2011). Auxine signalering: van synthese tot systeembiologie; een Subject collectie vanuit Cold Spring Harbor perspectieven in de biologie. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Google Scholar
Guilfoyle, T. J. (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar