Introdução
ao contrário dos animais, que pode mover-se para evitar o adversos ambiente, o sésseis plantas apresentam um altamente desenvolvido adaptação para as complicadas condições ambientais. Para alcançar esta profunda adaptabilidade, a comunicação entre as células é necessária. A comunicação célula a célula nas plantas envolve o processamento intracelular robusto de sinalização e redes intercelulares de sinalização intrincadas., Até agora, há pelo menos nove de sinalização de substâncias, denominada fábrica de hormônios, incluindo a auxina, brassinosteroid (BR), cytokinin, giberelinas (GA), etileno, jasmonic ácido (JA), strigolactone (SL), abscisic acid (ABA), e ácido salicílico (SA) descoberto (Druege et al., 2016; Verma et al., 2016). Os estudos genéticos e fisiológicos revelaram os papéis críticos e os mecanismos funcionais destes hormônios acima no crescimento e desenvolvimento das plantas (Gray, 2004)., Com base nos estudos anteriores, auxin, BR, GA, SL e citokinina funcionam principalmente durante o crescimento e desenvolvimento normal das plantas, enquanto ABA, etileno, JA e SA desempenham papéis importantes na resposta ao crescimento das plantas a vários estresses bióticos e abióticos (Pieterse et al., 2009; Santner et al., 2009; Denance et al., 2013). E também alguns desses hormônios têm papéis duplos, por exemplo, a ABA também desempenha papéis importantes no desenvolvimento de sementes e dormência (Seo e Koshiba, 2002)., Embora cada hormona desempenhe um papel predominante em certos aspectos, muitas hormonas sobrepuseram-se e as interacções das diferentes hormonas controlam muitos aspectos do desenvolvimento e do crescimento em resposta a sugestões endógenas de desenvolvimento e exógenas.a auxina e a BR são duas classes principais de hormonas promotoras do crescimento., BR, Um grupo de hormônios esteróides específicos da planta que poderiam interagir com outras fitohormonas como auxina, citoquinina, etileno, GA, JA e SA e regular uma ampla gama de processos de crescimento e desenvolvimento vegetal, incluindo germinação de sementes, alongamento celular, diferenciação vascular, formação e movimento estomata, floração e fertilidade masculina (Saini et al., 2015). Curiosamente, cada um destes processos também é controlado pelo auxin, sugerindo que estes dois hormônios interagem para controlar o desenvolvimento da planta., Nesta revisão, vamos delinear a transdução de sinal de auxin e BR com base no progresso recente e rever o crosstalk entre auxin e BR mediado crescimento e desenvolvimento da planta.
Auxin via sinalizadora
Auxin foi reconhecida pela primeira vez como hormônio vegetal devido ao seu papel no tropismo vegetal para a gravidade ou estímulos de luz., Mais tarde, o auxin foi identificado quimicamente como ácido indole-3-acético e mostrou desempenhar papéis essenciais na pletora de processos fisiológicos e de desenvolvimento vegetal, incluindo embriogénese, organogénese, diferenciação vascular, desenvolvimento de raízes e rebentos, crescimento trópico e desenvolvimento de frutas (Estelle, 2011).usando a análise genética em Arabidopsis, o mecanismo molecular subjacente à transdução do sinal de auxina foi bem investigado. Resposta aos inibidores de transporte 1 (TIR1) foi o primeiro receptor nuclear identificado do auxin (Ruegger et al., 1998; Dharmasiri et al., 2005)., TIR1 codifica uma proteína nuclear pertencente à proteína f-box como uma subunidade do complexo proteico de ubiquitina ligase SCF E3 (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005), para além da TIR1, existem três proteínas F-box adicionais, a saber, as proteínas AUXIN sinalizando F BOX (AFBs), que mostram actividade de ligação do auxin e mediam a sinalização do auxin em Arabidopsis (Badescu e Napier, 2006). O receptor TIR1 pode interagir com um grupo de proteínas AUX/IAA (ácido auxin/indole-3-acético) (Dharmasiri et al., 2003)., As proteínas AUX / IAA são reguladores negativos da sinalização auxina e há 29 membros do AUX/IAA codificados no genoma Arabidopsis. As proteínas do AUX / IAA podem interagir com a classe dos reguladores transcritionais, os factores de resposta do auxin (ARF), para mediar as respostas transcritionais ao auxin. Sob alto nível de auxina, as proteínas AUX / IAA interagem com a TIR1 como coreceptor da auxina, e podem ser ubiquitinadas pelo complexo SCFtir1, sendo assim degradadas através da via proteosoma ubiquitina (Gray et al., 2001; Lanza et al., 2012)., Após a destruição de AUX/IAA repressors, a auxina reguladores de transcrição ARFs que incluem 23 associações podem ser lançado a partir de AUX/IAA repressão e, assim, mediar a auxina respostas de ativação ou repressão de genes alvo (Guilfoyle e Hagen, 2007). Os diferentes conjuntos de proteína f-box e AUX / IAA ou ARFs inferem a complexidade durante a transdução do sinal auxin (Goh et al., 2012; Guilfoyle, 2015; Salehin et al., 2015).,
a acção coordenada dos repressores Transcritionais Aux / IAA e dos factores de transcrição ARF produz redes complexas de regulação genética que também foram notificadas em Physcomitrella (Lavy et al., 2016). Recentemente, descobriu-se que a subunidade CULLIN1 (CUL1) do SCF interage com a TIR1 e assim regula a estabilidade dos substratos da TIR1 e a sinalização auxin (Wang et al., 2016). A interacção entre TIR1 e Aux/IAA é também influenciada pela conformação espacial do Aux/IAAs, controlada por uma ciclofilina isomerase LRT2 em arroz (Jing et al., 2015)., O factor de choque térmico 90 (HSP90) e a co-acompanhante SGT1, respectivamente, interagem com a TIR1 e regula assim a estabilidade da TIR1, que afecta as interacções entre a TIR1 e a Aux/IAA e a sinalização da auxin (Wang et al., 2016).
além da via canónica canónica canónica-sinalizadora de auxina dependente da TIR1, a auxina tem sido recentemente relatada como provocando uma gama diversificada de respostas de desenvolvimento através de um mecanismo não canónico de sinalização de auxina., In this non-canonical auxin sensing process, ARF3 / ETTIN controls gene expression through interactions with process-specific transcription factors, which highly enriches auxin-mediated plant developmental diversity (Simonini et al., 2016, 2017).o BRASSINOSTERÓIDE foi descoberto pela primeira vez no pólen pela sua capacidade de promover o alongamento celular. Mais tarde, descobriu-se que a BR desempenha papéis em uma ampla gama de aspectos de crescimento de plantas e pode responder a tensões bióticas e abióticas., Hoje em dia a via de transdução do sinal BR foi amplamente clarificada por combinações de diferentes métodos, incluindo genética molecular, bioquímica, proteômica, e genômica, etc. A célula-superfície quinase BRASSINOSTEROID INSENSITIVE1 (BRI1) foi identificado como o receptor da BR que podem se ligar ao domínio extracelular de BRI1 e ativar sua atividade quinase e, assim, mudar num sinal de cascata para regular a transcrição (Li e Chory, 1997; Wang et al., 2001; Kinoshita et al., 2005; Kim and Wang, 2010; Clouse, 2011; Hothorn et al., 2011; She et al., 2011; Oh et al., 2012)., Após a percepção da BR, BRI1 interage com o co-receptor BRI1-associado à quinase 1 (BAK1) e suas homolog SOMATIC EMBRYOGENESISRECEPTOR KINASEs (SERKs) para formar um complexo receptor BR mais ativo (Li et al., 2002; Nam and Li, 2002; Wang et al., 2005; Tang et al., 2008; Gou et al., 2012). Fosforilatos BRI1 activados dois substratos de cinases citoplásmicas ancoradas com membranas de plasma: cinas sinalizadoras de BRASSINOSTERÓIDES (BSK1) e CRESCIMENTO1 diferencial constitutivo (CDG1) (Tang et al., 2008; Kim et al.,, 2011), que por sua vez fosforila uma fosfatase do tipo PP1 chamada BRI1-SUPRESSOR1 (BSU1) para ativar BSU1, levando à desosforilação BSU1 e inactivação da cinase brassinosteroid INSENSIBILA2 (BIN2). A atividade cinase do BIN2 também é inibida pela HISTONE DEACETILASE HDA6, que interage e desacetilatos no K189 do BIN2., Quando BR níveis são baixos, BRI1 está inativo devido a sua negativo regulador, BRI1 INIBIDOR de QUINASE 1 (BKI1) e proteína fosfatase 2A (PP2A), enquanto BIN2 phosphorylate dois BR homólogo fatores de transcrição, BRASSINAZOLE RESISTANT1 (BZR1) e BZR2 (também chamado de BES1 para BRI1-EMS-SUPRESSOR 1) (He et al., 2002; Wang et al., 2002; Yin et al., 2002; Mora-Garcia et al., 2004; Kim et al., 2009, 2011; Kim and Wang, 2010). Quando os níveis de BR são altos, BIN2 é inactivado, e BZR1 e BZR2 são desphosporilados por PP2A, e movem-se para o núcleo para alterar a expressão de milhares de genes de resposta BR (He et al.,, 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011).
A Sinergia Entre a BR e a Auxina Sinalização
Auxina e BR vias de sinalização desempenhar diversos papéis, no entanto, eles também mostraram sinérgica e interdependentes medicamentosas em uma ampla gama de processos de desenvolvimento. Por exemplo, sinais auxin e BR podem promover a expansão celular e interagir sinergisticamente para promover o alongamento hipocotílico (Nemhauser et al., 2004)., A resposta de uma das duas vias na promoção do alongamento hipocotílico requer a função da outra e a interdependência entre as vias BR e auxin (Nemhauser et al., 2004). A auxina aumentou o comprimento hipocotílico em plantas de tipo selvagem, mas não no bri1-116 mutante insensível à BR, e este fenótipo insensível à auxina do bri1-116 foi suprimido pelo bzr1-1D mutante com ganho de função dominante, indicando BR ou bzr1 activo é necessário para a promoção de Elongação hipocotílica pela auxina., Descobriu-se que a sinalização BR converge com o supressor do Fitocromo B4-3 (SOB3) para controlar a elongação celular e o crescimento hipocotílico através da regulação da auxina induzida pela pequena expressão RNA19 (SAUR19) (Favero et al., 2017). Por outro lado, a auxina regulamentado fator de transcrição ÓRGÃO PEQUENO TAMANHO 1 (SMOS1) foi recentemente encontrada para controlar a expansão celular através da interação direta com SMOS2/DLT, um membro do GRAS família de transcrição co-reguladores, que desempenha um papel positivo na BR sinalização no arroz (Kim et al., 2009; Tong et al., 2012; Hirano et al., 2017)., Auxina relacionados mutantes, tais como iaa3 e arf6/arf8 foram menos sensíveis à BR que foi de tipo selvagem para hypocotyl alongamento, e aboliu a hipersensibilidade do bzr1-1D para auxina, sugerindo a BR e BZR1 promoção de hypocotyl alongamento requer ARF6/8. A análise do ChIP-Seq revelou que o ARF6 compartilha um grande número de alvos genômicos (cerca de 50%) com o BZR1 e o Fator de transcrição regulado pela luz/temperatura PIF4 por análises CHIP-Seq (Oh et al., 2014)., BZR1 e PIF4 interagem com ARF6 e ativam genes alvo compartilhados ligando-se aos genes alvo compartilhados cooperativamente durante o alongamento hipocotílico (Oh et al., 2014) e muitos desses genes-alvo sobrepostos codificam proteínas da parede celular envolvidas na expansão celular.Brassinosteroid and auxin also play important roles in the maintenance of root apical meristem (RAM) (Durbak et al., 2012). A RAM consiste de um pequeno grupo de células raramente dividindo conhecidas como o centro quiescente (CQ), cercadas por células-tronco que dão origem aos vários tipos de tecidos toot., A manutenção da população de células estaminais da raiz é regulada pela Homebox 5 (Wox5) (Sarkar et al., 2007). O WOX5 é restrito ao CQ por sinalização auxin e facilita a expressão adequada dos genes PLT (Aida et al., 2004; Ding and Friml, 2010). Mutações no gene receptor BR BRASSINOSTEROID insensível 1 (BRI1) resultam na progressão do ciclo celular aberrante na RAM e causam um RAM menor (Gonzalez-Garcia et al., 2011; Hacham et al., 2011). O Auxin é conhecido por estimular a biossíntese da BR (Chung et al.,, 2011), mas a atividade da BR não afeta a expressão de genes PIN (Hacham et al., 2011). Os fenótipos da ponta da raiz dos mutantes BR não mostram o mesmo que os mutantes auxin (Gonzalez-Garcia et al., 2011), indicando que a BR age sobre a RAM independentemente do auxin.os sinais de Brassinosteróide e de auxina também são sinergisticamente necessários na formação de padrões radiais de feixes vasculares (Ibanes et al., 2009)., Pelas combinações de modelagem matemática e experimentos biológicos, auxin maxima, estabelecido pelo transporte polar auxin assimétrico, mas não mudanças nos níveis de auxin é importante para posicionar os feixes vasculares. O sinal BR foi mostrado para servir como um sinal de promoção para o número de células no anel provascular que são consistentes com auxin maxima. Assim, o estabelecimento de um arranjo periódico de feixes vasculares no disparo está sob a ação coordenada destes dois hormônios vegetais (Ibanes et al., 2009)., Ambos os sinais também estão envolvidos no desenvolvimento da raiz vegetal e a interação de BR e auxin é mediada por BREVIS RADIX (BRX) durante este processo. A BRX é importante para a biossíntese limitante da taxa de BR e da aplicação exógena BR pode salvar defeitos mutantes brx. Além disso, a expressão genética do auxin é globalmente prejudicada no mutante brx, e a expressão do BRX é fortemente induzida pelo auxin e suprimida pelo BR, implicando biossíntese BR e sinalização auxin são conectados através de um ciclo de feedback envolvendo BRX durante o desenvolvimento da raiz (Mouchel et al., 2006).,os Brassinosteróides e a auxina também desempenham funções sinérgicas durante o desenvolvimento lateral da raiz. As BRs funcionam principalmente na iniciação da raiz lateral primordia, enquanto o auxin é necessário tanto para os estágios de iniciação como de emergência da formação da raiz lateral (Casimiro et al., 2001; Bhalerao et al., 2002; Benkova et al., 2003; Bao et al., 2004). Durante estes processos, a BRs aumenta a iniciação LRP promovendo o transporte de auxina acropetal na raiz, mas não afetando o nível IAA endógeno (Bao et al., 2004)., Todos estes relatórios sugerem que o crosstalk entre a BR e o auxin desempenha um papel importante no crescimento e desenvolvimento das instalações de regulação.
br Regulates Auxin Signaling
aparte The interdependency and cooperation of auxin and BR signals during plant development, BR could mediate auxin signal pathway on multiple levels. O BZR1 interage com proteínas ARF para atingir diretamente múltiplos componentes sinalizadores de auxina e genes envolvidos no metabolismo do auxin, tais como transporte e sinalização, incluindo AUX/IAA, PINs, TIR1, e ARFs, etc. (Sun et al., 2010)., Verificou-se que as proteínas Aux/IAA estão envolvidas nas respostas BR e que os mutantes Iaa7/axr2-1 e iaa17/axr3-3 mostraram sensibilidade BR aberrante e expressão genética induzida por br aberrante de uma forma dependente de órgãos (Nakamura et al., 2006). O tratamento com brassinolida exógena (BL) poderia induzir a expressão de genes que respondiam às auxinas, tais como IAA5, IAA19, IAA17, etc. e a expressão acima genes é reduzida na BR biossintéticos mutante de-etiolated2 (det2), o que indica que o funcional BR biossíntese é, em parte, necessária para a auxina dependente da expressão do gene (Nakamura et al.,, 2003; Kim et al., 2006). Adicionalmente, a BR também afeta o fluxo de auxin regulando a expressão de exportadores de auxin como PIN4 e PIN7 (Nakamura et al., 2004). Durante a planta gravitropism respostas, BRs poderia melhorar o polar acúmulo de auxina exportador PIN2 na raiz meristem zona e, portanto, afetam a redistribuição de auxina da raiz a ponta em direção ao alongamento e zonas de resultar na diferença de IAA níveis em ambos os lados superior e inferior das raízes para induzir a planta gravitropism., Durante este processo, o ROP2 ativado pela BR desempenha um papel importante na modulação da localização funcional do PIN2 através da regulação da montagem/remontagem de f-actins (Li et al., 2005). Estudos adicionais mostraram que a diminuição da percepção e/ou concentração de BL pode induzir o CYP79B2, o gene que codifica uma enzima que converte o triptofano em indole-3-acetaldoxima e, assim, afectar a distribuição (Kim et al., 2007).
In addition, it was found that BR signal could regulate auxin signaling output by its negative regulator GSK3 kinase BIN2., O factor de Resposta da auxina, o ARF2, foi identificado como uma proteína interagindo com o BIN2 num ecrã híbrido de levedura e o ensaio de kinase mostrou que o BIN2 podia fosforilar o ARF2. A fosforilação da ARF2 resulta na perda da sua capacidade de ligação ao ADN e da actividade de repressão dos genes-alvo (Vert et al., 2008). ARF2 é um gene alvo BZR1 e sua expressão é reduzida pelo tratamento BR (Sun et al., 2010)., Além disso, BIN2 pode phosphorylate ARF7 e ARF19 para suprimir sua interação com o AUX/IAAs e, assim, melhorar a atividade transcricional em seus genes alvo LATERAL ÓRGÃO LIMITES-DOMAIN16 (LBD16) e LBD29 para regular lateral de raiz organogénese (Cho et al., 2014). No entanto, BR desempenha um papel menor durante este processo e BIN2 está sob o controle do fator inibitório de diferenciação do elemento TRAQUEÁRIO (TDIF)–módulo RECEPTOR TDIF (TDR) (Cho et al., 2014). Juntos, a BR pode regular reponses auxin influenciando diferentes componentes de sinalização auxin.,
Auxin Regulates BR Signaling
On the other hand, auxin can also regulate BR signal pathway in certain aspects. A expressão de DWARF4, uma hidroxilase crucial para a biossíntese BR para controlar o nível de BR endógeno, é dependente da auxina. O tratamento com Auxin poderia estimular visivelmente a expressão de DWARF4 e o auxin poderia inibir a ligação de BZR1 ao promotor de DWARF4. A indução da DWARF4 pelo auxin requer a via sinalizadora do auxin, mas não a via sinalizadora BR (Chung et al., 2011; Yoshimitsu et al., 2011)., A oxidação C-3 catalisadora da BR foi ativada pela BRX, um fator de transcrição putativo atuando a jusante da sinalização auxina (Mouchel et al., 2006). Estudos adicionais em rice indicam que a auxina exógena pode melhorar os níveis de expressão de transcrição do gene receptor BR OsBRI1, sugerindo que a auxina aumenta a sinalização BR através da regulação dos receptores BR (Sakamoto et al., 2013). Além disso, o promotor da OsBRI1 possui um motivo de auxina-resposta a montante (AuxRE) que é alvo de factores de transcrição ARF., Além disso, estudos mutantes indicam que após a mutação de AuxRE, a indução de expressão de OsBRI1 por auxin é abolida e também a expressão de OsBRI1 é regulada para baixo em mutante de arf (Sakamoto et al., 2013). It has been reported that OsARF19 binds to the promoter of OsBRI1 and positively regulates its expression which then activates the BR signaling (Zhang et al., 2015). O BES1 pode ligar-se ao promotor do pequeno RNA AUXIN-UP 15 (SAUR15) e mediar o gene de resposta precoce BR em Arabidopsis, e esta ligação pode ser reforçada pelo tratamento com auxin (Walcher e Nemhauser, 2012)., Em conjunto, o auxin também pode afetar as respostas BR e o crescimento e desenvolvimento regulados da planta BR.durante as últimas quase quatro décadas, estudos sobre interações de vias Auxin-BR atraíram cada vez mais o interesse dos investigadores. O aparelho de ferramentas fisiológicas, moleculares, genéticas e bioquímicas aprofundaram muito a nossa compreensão desta questão., Com base nos estudos anteriores, BR e auxin estão envolvidos sinergisticamente em múltiplos processos de desenvolvimento vegetal, incluindo: alongamento hipocotílico, desenvolvimento de feixes vasculares, desenvolvimento de raízes e tropismos, etc. A interdependência e cooperação do auxin e da BR são complicadas e envolvem numerosos processos a nível molecular, partilhando os mesmos genes-alvo, regulando-se mutuamente em vários níveis (Figura 1).
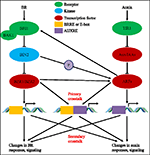
a FIGURA 1. Modelo de auxin-brassinosteroid (BR) crosstalk., Em Arabidopsis, as percepções do sinal BR e auxin são reconhecidas pelos receptores BRI1 e TIR1, respectivamente. A BR liga-se ao domínio extracelular da BRI1 e promove a interacção com o co-receptor BAK1 para formar um complexo de receptores BR mais activo, o que por sua vez leva à desorganização e inactivação da BIN2. A inactivação do BIN2 leva à desphosporilação de dois factores de transcrição homólogos BZR1 e BZR2, que se deslocam para o núcleo para activar a transcrição de genes que contêm BRRE ou caixa electrónica na sua região promotora., BIN2 também pode fosforilar ARF7 e ARF19 para suprimir sua interação com AUX/IAAs e, assim, aumentar a atividade transcritional em seus genes alvo. Receber o sinal de auxina e interagir com as proteínas AUX/IAA como co-receptor da auxina. O AUX/IAA, em seguida, é degradada através de hidrolase–proteassoma caminho, e a auxina transcricional reguladores de auxina fatores de resposta (ARFs) são liberados a partir AUX/IAA repressão e ativar a transcrição de genes com auxina elementos sensíveis (AUXRE) na respectiva regulamentação região., Algumas ARFs também podem se ligar ao promotor de BRI1 e regula positivamente sua expressão, que então ativa a sinalização BR. O crosstalk primário ocorre pela ativação de genes que contêm tanto BRRE/e-box quanto AUXRE em sua região promotora, permitindo que ambas as vias de sinalização regulem diretamente a transcrição. Crosstalk secundário ocorre através da expressão de genes que são auxin ou BR responsivos, mas as atividades das quais controlam a expressão de genes que regulam a resposta e sinalização de outros hormônios.,
Fosforilação regulamento desempenha um papel crucial na BR via de sinalização, especialmente durante o processo de percepção, BR é percebido através BRI1 quinase do receptor e BAK1 quinase co-receptores, e, eventualmente, controles BR regulamentado expressão de genes influenciando a jusante fatores de transcrição, tais como BES1/BZR1 atividades (He et al., 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011). No entanto, a regulação da ubiquitinação parece essencial para a sinalização auxina., Uma vez auxina liga TIR1 receptor, que atua como uma hidrolase E3-ligase, ativado TIR1 E3-ligase ubiquitinates AUX/IAA proteínas, leva à degradação destes repressors e de represses ARF fatores de transcrição, e, eventualmente, faz com auxina regulamentado padrão de expressão do gene mudanças e crescimento respostas (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005). Uma vez que se descobriu que a BIN2 kinase, que é bem conhecida por funcionar na sinalização BR, poderia fosforilar e melhorar as atividades de ARFs, como ARF2 e ARF7 (Vert et al., 2008; Cho et al.,, 2014), será interessante testar se cinases como BIN2, que estão envolvidas na sinalização BR, também podem interagir com outros componentes de sinalização auxin, como receptores TIR1 ou repressores AUX/IAA, e influenciar a atividade TIR1 E3-ligase ou estabilidades proteicas AUX/IAA. Por outro lado, o papel da ubiquitinação na sinalização BR também precisa ser abordado, especialmente se TIR1 E3-ligase poderia interagir diretamente com os componentes de sinalização BR e regular suas estabilidades proteicas.,
além disso, o uso auxina resposta DR5 e outros auxina repórteres, tem sido observado que a auxina regula o crescimento e desenvolvimento das plantas em um tecido ou celular dependente maneira. As diversas saídas transcritionais dependendo do contexto celular e ambiental (Clark et al., 2014; Etchells et al., 2016; Lavy et al., 2016)., Embora o spatiotemporal BR sinalização tem sido mostrado para controlar o crescimento da raiz, através da ação antagônica com auxina (Chaiwanon e Wang, 2015), ainda é desconhecido se o tecido ou celular BR sinalização, o que pode ser visualizado pelo pBZR1:BZR1-YFP, também é importante para o controle de outros processos além do desenvolvimento do sistema radicular. Além disso, a geração de um mapa detalhado dos tecidos ou adega das distribuições de auxina e BR é atualmente possível utilizando a triagem celular activada por fluorescência ou Microdissecção por laser em combinação com a análise da expressão genética de alta resolução., Isto acabará por levar ao endereço se o auxin crosstalks Com BR em um tecido ou maneira celular.
contribuições dos autores
todos os autores estiveram envolvidos na escrita deste manuscrito de revisão. Para mais informações sobre o que constitui autoria, por favor consulte as nossas Diretrizes de Autor.,Este trabalho é apoiado por subvenções da Fundação Nacional de Ciência Natural da China (projetos 31500229 e 31770305), da Fundação de Ciência Natural da província de Shandong para o principal programa de pesquisa básica (2017C03), fundos provinciais de Shandong para jovens acadêmicos ilustres (2014JQ201408).
Declaração de conflito de interesses
os autores declaram que a investigação foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.
Clouse, S. D. (2011)., Transdução do sinal de brassinosteróide: desde a activação da cinase receptora até às redes de transcrição que regulam o desenvolvimento das instalações. Plant Cell 23, 1219-1230. doi: 10.1105/tpc.111.084475
PubMed Abstract | CrossRef Full Text/Google Scholar
Estelle, M. (2011). Auxin Signaling: From Synthesis to Systems Biology; A Subject Collection from Cold Spring Harbor Perspectives in Biology. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Google Scholar
Guilfoyle, T. J. (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar