Introducere
spre deosebire de animalele care se pot deplasa pentru a evita împrejurimile adverse, plantele sesile prezintă o adaptare foarte dezvoltată la condițiile complicate de mediu. Pentru a realiza această adaptabilitate profundă, sunt necesare comunicări între celule. Comunicarea celulă-celulă în plante implică o prelucrare robustă a semnalizării intracelulare și rețele complicate de semnalizare intercelulară., Până acum, există cel puțin nouă semnalizare substanțe, numite hormoni de plante, inclusiv auxina, brassinosteroid (BR), citochinine, gibereline (GA), etilenă, jasmonic acid (JA), strigolactone (SL), acid abscisic (ABA), și acid salicilic (SA) a descoperit (Druege et al., 2016; Verma și colab., 2016). Studiile genetice și fiziologice au relevat rolurile critice și mecanismele funcționale ale acestor hormoni de mai sus în creșterea și dezvoltarea plantelor (Gray, 2004)., Pe baza studiilor anterioare, auxin, BR, GA, SL și cytokinin funcționează în principal în timpul creșterii și dezvoltării normale a plantelor, în timp ce ABA, etilena, JA și SA joacă roluri importante în răspunsul creșterii plantelor la diferite stresuri biotice și abiotice (Pieterse et al., 2009; Santner și colab., 2009; Denance și colab., 2013). Și, de asemenea, unii dintre acești hormoni au roluri duble, de exemplu, ABA joacă, de asemenea, roluri importante în dezvoltarea semințelor și dormirea (Seo și Koshiba, 2002)., Deși fiecare hormon joacă roluri predominante în anumite aspecte, mulți hormoni au activități suprapuse și interacțiunile diferiților hormoni controlează multe aspecte de dezvoltare și creștere ca răspuns la indiciile endogene de dezvoltare și exogene.Auxin și BR sunt două clase majore de hormoni care promovează creșterea., BR, un grup de plante specifice hormoni steroizi care ar putea interacționa cu alți fitohormoni, cum ar fi auxina, citochinine, etilenă, GA, DA, si SA și de a reglementa o gama larga de creștere a plantelor și procese de dezvoltare, inclusiv de germinare a semințelor, alungirea celulelor, vasculare diferențiere, stomate formarea și mișcarea, înflorire și fertilitatea masculină (Saini et al., 2015). Interesant este că fiecare dintre aceste procese este controlat și de auxin, sugerând interacțiunea acestor doi hormoni pentru a controla dezvoltarea plantelor., În această revizuire, vom sublinia transducția semnalului auxin și BR pe baza progreselor recente și vom revizui discuția dintre creșterea și dezvoltarea plantelor mediate de auxin și BR.Auxin a fost recunoscut pentru prima dată ca hormon de plante datorită rolului său în tropismul plantelor la gravitație sau stimuli ușori., Mai târziu auxina a fost din punct de vedere chimic identificat ca indol-3-acetic acid și prezentate pentru a juca roluri esențiale în multitudinea de plante de dezvoltare și procese fiziologice, inclusiv embriogeneza, organogeneză, vasculare diferențiere, rădăcină și trage de dezvoltare, tropic creșterea și dezvoltarea fructelor (Estelle, 2011).folosind analiza genetică în Arabidopsis, mecanismul molecular care stă la baza transducției semnalului auxin a fost bine investigat. Răspunsul inhibitorului de TRANSPORT1 (TIR1) a fost primul receptor nuclear identificat al auxinei (Ruegger et al., 1998; Dharmasiri și colab., 2005)., TIR1 codifică o proteină nucleară aparținând proteinei F-box ca o subunitate a complexului proteic de ligază ubiquitin SCF E3 (Gray et al., 1999, 2002; Hellmann și colab., 2003; Quint și colab., 2005), în plus față de TIR1, există trei proteine suplimentare F-box și anume proteinele de semnalizare auxină F BOX (AFBs) care prezintă activitate de legare a auxinei și mediază semnalizarea auxinei în Arabidopsis (Badescu și Napier, 2006). Receptorul TIR1 poate interacționa cu un grup de proteine aux/IAA (auxin/indol-3-acid acetic) (Dharmasiri și colab., 2003)., Proteinele aux/IAA sunt regulatori negativi ai semnalizării auxinelor și există 29 de membri ai aux / IAA codificați în genomul Arabidopsis. Proteinele aux / IAA ar putea interacționa cu clasa regulatorilor transcripționali, factorii de răspuns auxin (ARF), pentru a media răspunsurile transcripționale la auxin. În mare auxin nivel, AUX/IAA proteine interacționează cu TIR1 ca coreceptor de auxina, și poate fi ubiquitinated de SCFtir1 complex și, astfel, să fie degradat prin ubiquitin–proteazomal cale (Gray et al., 2001; Lanza și colab., 2012)., După distrugerea represorilor aux / IAA, regulatorii ARF de transcripție auxin care includ 23 de membri pot fi eliberați de represiunea aux / IAA și astfel pot Media răspunsurile auxin prin activarea sau reprimarea genelor țintă (Guilfoyle și Hagen, 2007). Diferitele seturi de proteine F-box și AUX / IAA sau ARFs deduc complexitatea în timpul transducției semnalului auxin (Goh et al., 2012; Guilfoyle, 2015; Salehin și colab., 2015).,
o acțiune coordonată de Aux/IAA transcripțional repressors și ARF factori de transcriere produce complex de gene-retele de reglementare care au fost raportate, de asemenea, în Physcomitrella (Lavy et al., 2016). Recent, s-a constatat că CULLIN1 (CUL1) subunitatea de CSA interacționează cu TIR1 și reglementează, astfel, TIR1 substraturi de stabilitate și de auxina de semnalizare (Wang et al., 2016). Interacțiunea dintre TIR1 și Aux/IAA este, de asemenea, influențată de conformație ale Aux/IAAs, controlat de un cyclophilin izomeraza LRT2 în orez (Jing et al., 2015)., ȘOC TERMIC FACTOR de 90 (HSP90) și co-însoțitor SGT1, respectiv, interacționează cu TIR1 și reglementează, astfel, TIR1 de stabilitate, care afectează interacțiunile dintre TIR1 și Aux/IAA și auxin de semnalizare (Wang et al., 2016).
pe lângă calea de semnalizare auxină canonică dependentă de TIR1, auxin a fost recent raportat că provoacă o gamă variată de răspunsuri de dezvoltare printr-un mecanism de semnalizare auxină non-canonic., În acest non-canonice auxin de detectare proces, ARF3/ETTIN controlează expresia genelor prin interacțiuni cu procesul specific de factori de transcripție, care îmbogățește foarte auxin-mediată de plante de dezvoltare diversitate (Simonini et al., 2016, 2017).
calea de semnalizare BR
BRASSINOSTEROID a fost descoperit pentru prima dată în polen pentru capacitatea sa de a promova alungirea celulelor. Mai târziu, sa constatat că BR joacă roluri într-o gamă largă de aspecte de creștere a plantelor și poate răspunde la stresul biotic și abiotic., În prezent, calea de transducție a semnalului BR A fost clarificată în mare măsură prin combinații de metode diferite, inclusiv genetică moleculară, biochimie, proteomică și genomică etc. Celula-suprafața kinaza BRASSINOSTEROID INSENSITIVE1 (BRI1) a fost identificat ca receptor de BR care se pot lega la domeniul extracelular al BRI1 și activarea kinazei și, astfel, a comuta la un semnal cascadă de a reglementa transcriere (Li și Chory, 1997; Wang et al., 2001; Kinoshita și colab., 2005; Kim și Wang, 2010; Clouse, 2011; Hothorn și colab., 2011; ea și colab., 2011; Oh și colab., 2012)., La percepția BR, BRI1 interacționează cu co-receptorilor BRI1 ASOCIATE KINAZA 1 (BAK1) și homolog SOMATICE EMBRYOGENESISRECEPTOR Kinaze (SERKs) pentru a forma o mai activă BR complex receptor (Li et al., 2002; Nam și Li, 2002; Wang și colab., 2005; Tang și colab., 2008; Gou și colab., 2012). Activat BRI1 phosphorylates două substraturi ale membranei plasmatice-ancorat receptor, cum ar fi kinaze citoplasmatice: BRASSINOSTEROID-SEMNALIZARE KINASES1 (BSK1) și CONSTITUTIV DIFERENȚIAL MUNCĂ1 (CDG1) (Tang et al., 2008; Kim și colab.,, 2011), care, la rândul său phosphorylates o PP1-tip fosfataza numit BRI1-SUPPRESSOR1 (BSU1) pentru a activa BSU1, ceea ce duce la BSU1 dephosphorylation și inactivarea de GSK3-ca kinaza BRASSINOSTEROID INSENSITIVE2 (BIN2). La activitatea kinazei de BIN2 este, de asemenea, inhibată de HISTONE DEACETYLASE HDA6, care interacționează și deacetylates la K189 de BIN2., Când BR nivelurile sunt scăzute, BRI1 este pasiv datorită reglator negativ, BRI1 INHIBITOR al KINAZEI 1 (BKI1) și proteine fosfataza 2A (pachetul de măsuri 2a), în timp ce BIN2 fosforila două BR omoloage factori de transcriere, BRASSINAZOLE RESISTANT1 (BZR1) și BZR2 (de asemenea, numit BES1 pentru BRI1-EMS-SUPRESOARE 1) litera (A et al., 2002; Wang și colab., 2002; Yin și colab., 2002; Mora-Garcia și colab., 2004; Kim și colab., 2009, 2011; Kim și Wang, 2010). Când BR nivelurile sunt ridicate, BIN2 este inactivat, și BZR1 și BZR2 sunt dephosphorylated de pachetul de măsuri 2a, și pentru a muta în nucleu să-și modifice expresia de mii de BR răspuns gene (A et al.,, 2005; Yin și colab., 2005; Sun et al., 2010; Tang și colab., 2011; Yu și colab., 2011).
sinergia dintre semnalizarea BR și Auxin
căile de semnal Auxin și BR joacă roluri diverse, cu toate acestea, au arătat, de asemenea, interacțiuni sinergice și interdependente într-o gamă largă de procese de dezvoltare. De exemplu, ambele semnale auxin și BR pot promova expansiunea celulelor și pot interacționa sinergic pentru a promova alungirea hipocotililor (Nemhauser et al., 2004)., Răspunsul uneia dintre cele două căi în promovarea alungirii hipocotil necesită funcția celeilalte și interdependența dintre căile BR și auxin (Nemhauser et al., 2004). Auxina crescut hypocotyl lungimea de tip sălbatic plante, dar nu în BR-insensibil mutant bri1-116, iar acest lucru auxin-insensibil fenotip de bri1-116, a fost suprimat prin dominant câștig de funcție mutant bzr1-1D, indicând BR sau active BZR1 este necesar pentru auxin promovarea hypocotyl alungire., S-a constatat că BR semnalizare converge cu SUPRESOARE DE PHYTOCHROME B4-3 (SOB3) pentru a controla alungirea celulelor și hipocotilul de creștere prin regulamentul de auxina induse MICI AUXIN PÂNĂ RNA19 (SAUR19) expresia (Favero et al., 2017). Pe de altă parte, auxin reglementate factor de transcriere ORGAN MIC, MARIMEA 1 (SMOS1) a fost recent descoperit la controlul de expansiune de celule prin interacțiune directă cu SMOS2/DLT, un membru al CARNAVALULUI familie de transcriptie co-reglementare, care joacă un rol pozitiv în BR semnalizare în orez (Kim et al., 2009; Tong și colab., 2012; Hirano și colab., 2017)., Auxina legate de mutanți, cum ar fi iaa3 și arf6/arf8 au fost mai puțin sensibile la BR decât era de tip sălbatic pentru hypocotyl alungire, și abolit de hipersensibilitate de bzr1-1D la auxina, sugerând BR BZR1 și promovarea hypocotyl alungirea necesită ARF6/8. La nivelul întregului genom ChIP-Seq analiza a arătat că ARF6 acțiuni un număr mare de genomică obiective (aproximativ 50%) cu BZR1 și lumina/temperatura reglementate factor de transcriere PIF4 de CHIP-Seq analize (Oh et al., 2014)., BZR1 și PIF4 interacționa cu ARF6 și de a activa obiectiv comun genelor prin legarea sa de obiectiv comun de gene cooperativ în timpul hypocotyls alungire (Oh et al., 2014) și multe dintre aceste gene țintă care se suprapun codifică proteinele peretelui celular implicate în expansiunea celulară.Brassinosteroid și auxin joacă, de asemenea, roluri importante în menținerea meristemului apical rădăcină (RAM) (Durbak și colab., 2012). Berbecul constă dintr-un grup mic de celule care se divid rar, cunoscut sub numele de centrul de repaus (QC), înconjurat de celule stem care dau naștere diferitelor tipuri de țesuturi., Menținerea populației de celule stem rădăcină este reglementată de HOMEOBOX 5 (WOX5) legat de WUSCHEL (Sarkar et al., 2007). WOX5 este limitat la QC prin semnalizarea auxinei și facilitează exprimarea corectă a genelor PLT (Aida et al., 2004; Ding și Friml, 2010). Mutațiile genei receptorului BR BRASSINOSTEROID insensibil 1 (BRI1) au ca rezultat progresia aberantă a ciclului celular în Berbec și provoacă o RAM mai mică (Gonzalez-Garcia et al., 2011; Hacham și colab., 2011). Auxin este cunoscut pentru a stimula biosinteza BR (Chung și colab.,, 2011), dar activitatea BR nu afectează expresia genelor PIN (Hacham și colab., 2011). Fenotipurile vârfului rădăcinii mutanților BR nu arată la fel ca mutanții auxin (Gonzalez-Garcia et al., 2011), indicând faptul că BR acționează asupra RAM independent de auxin.semnalele Brassinosteroid și auxin sunt, de asemenea, necesare sinergic în formarea modelului radial al fasciculelor vasculare (Ibanes și colab., 2009)., Prin combinațiile de modelare matematică și experimente biologice, auxin maxima, stabilită prin transportul polar auxin asimetric, dar nu și modificările nivelurilor auxin este importantă pentru poziționarea fasciculelor vasculare. Semnalul BR sa dovedit a servi ca un semnal de promovare a numărului de celule din inelul provascular care sunt în concordanță cu auxin maxima. Astfel, stabilirea aranjamentului periodic al fasciculelor vasculare în tragere este sub acțiunea coordonată a acestor doi hormoni de plante (Ibanes et al., 2009)., Ambele semnale sunt, de asemenea, implicate în dezvoltarea rădăcinilor plantelor, iar interacțiunea dintre BR și auxin este mediată de BREVIS RADIX (BRX) în timpul acestui proces. BRX este important pentru biosinteza limitatoare de viteză a BR și BR aplicarea exogenă poate salva defectele mutante brx. În plus, auxin-receptiv expresia genică la nivel global este afectată în brx mutant, și expresia de BRX este puternic induse de auxina și suprimat de BR, implicând BR biosinteza și auxin de semnalizare sunt conectate printr-o bucla de feedback care implică BRX timpul rădăcină de dezvoltare (Mouchel et al., 2006).,Brassinosteroidele și auxina joacă, de asemenea, roluri sinergice în timpul dezvoltării rădăcinilor laterale. BRs funcționează în principal la inițierea primordiei radiculare laterale, în timp ce auxina este necesară atât pentru etapele de inițiere, cât și pentru apariția formării radiculare laterale (Casimiro et al., 2001; Bhalerao și colab., 2002; Benkova și colab., 2003; Bao și colab., 2004). În timpul acestor procese, BRs crește inițierea LRP prin promovarea transportului de auxine acropetale în rădăcină, dar nu prin afectarea nivelului IAA endogen (Bao et al., 2004)., Toate aceste rapoarte sugerează că discuția dintre BR și auxin joacă un rol important în creșterea și dezvoltarea plantelor de reglementare.pe lângă interdependența și cooperarea semnalelor auxin și BR în timpul dezvoltării plantelor, BR ar putea media calea semnalului auxin pe mai multe niveluri. BZR1 interacționează cu ARF proteine să vizeze în mod direct mai multe auxin semnalizare componente și gene implicate în metabolismul auxinelor, cum ar fi transport și de semnalizare, inclusiv AUX/IAA, Ace, TIR1, și Arf, etc. (Sun și colab., 2010)., S-a constatat că Aux/IAA proteine sunt implicate în BR răspunsuri și iaa7/axr2-1 și iaa17/axr3-3 mutanți arătat aberante BR sensibilitate și aberant BR-a indus expresia genelor într-un organ manieră dependentă (Nakamura et al., 2006). Exogene brassinolide (BL) tratament ar putea induce expresia auxin-receptiv gene, cum ar fi IAA5, IAA19, IAA17, etc. și expresia de mai sus de gene este în jos-reglementată în BR biosinteză mutant de-etiolated2 (det2), care indică faptul că funcționale BR biosinteza este parțial necesare pentru auxin-dependente expresia genelor (Nakamura et al.,, 2003; Kim și colab., 2006). În plus, BR afectează, de asemenea, fluxul de auxină prin reglarea expresiei exportatorilor de auxină, cum ar fi PIN4 și PIN7 (Nakamura et al., 2004). Timpul de plante gravitropism răspunsuri, BRs ar putea spori polar acumularea auxinei exportator PIN2 în rădăcina de meristeme zonă și, astfel, afecta redistribuire a auxina din rădăcină vârful spre zonele de alungire și de rezultat din diferența de IAA niveluri în ambele părți superioare și inferioare de rădăcini pentru a induce plantelor gravitropism., În timpul acestui proces, ROP2 activat BR joacă un rol important în modularea localizării funcționale a PIN2 prin reglarea asamblării / reasamblării F-actinelor (Li et al., 2005). Studii suplimentare au arătat că a scăzut BL percepția și/sau de concentrare ar putea induce CYP79B2, gena care codifică o enzimă de conversie triptofan la indol-3-acetaldoxime și, astfel, afectează distribuția (Kim et al., 2007).în plus ,sa constatat că semnalul BR ar putea regla ieșirea de semnalizare auxin prin regulatorul său negativ gsk3 kinase BIN2., Auxinei factorul de răspuns ARF2 a fost identificat ca un BIN2 interacționează proteine într-o drojdie de două-hibrid ecran și kinaza testul a arătat BIN2 poate fosforila ARF2. Fosforilarea ARF2 are ca rezultat pierderea capacității sale de legare a ADN-ului și a activității de represiune a genelor țintă (Vert et al., 2008). Arf2 este un gene țintă BZR1 și expresia sa este redusă prin tratamentul BR (Sun et al ., 2010)., În plus, BIN2 poate fosforila ARF7 și ARF19 pentru a suprima interacțiunea lor cu AUX/IAAs și, astfel, a spori activitatea transcripțională pe gene tinta LATERALE ORGAN LIMITE-DOMAIN16 (LBD16) și LBD29 de a reglementa laterale rădăcină de organogeneză (Cho et al., 2014). Cu toate acestea, BR joacă un rol minor în timpul acestui proces, și BIN2 este sub controlul TRACHEARY ELEMENT de DIFERENȚIERE INHIBITOR FACTOR (TDIF)–TDIF RECEPTORILOR (TDR) modulul (Cho et al., 2014). Împreună, BR poate reglementa reponses auxin prin influențarea diferitelor componente de semnalizare auxin.,pe de altă parte, auxin poate regla, de asemenea, calea semnalului BR În anumite aspecte. Expresia DWARF4, o hidroxilază crucială pentru biosinteza BR pentru a controla nivelul BR endogen, este dependentă de auxină. Auxina tratament ar putea vizibil stimula expresia DWARF4 și auxin-ar putea inhiba legarea de BZR1 la promotor de DWARF4. Inducerea DWARF4 de către auxin necesită calea de semnalizare auxin, dar nu calea de semnalizare BR (Chung și colab., 2011; Yoshimitsu și colab., 2011)., CPD catalizând oxidarea C-3 A BR A fost activată de BRX, un factor de transcripție presupus care acționează în aval de semnalizarea auxinei (Mouchel et al., 2006). Studiul suplimentar în orez indică faptul că auxina exogenă poate spori nivelurile de expresie a transcripției genei receptorului br OsBRI1, sugerând că auxina îmbunătățește semnalizarea BR prin reglarea receptorilor BR (Sakamoto et al., 2013). Mai mult, promotorul OsBRI1 posedă un motiv element auxin-răspuns în amonte (AuxRE) care este vizat de factorii de transcripție ARF., Mai mult decât atât, mutant studii indică faptul că la o mutație de AuxRE, inducerea de exprimare a OsBRI1 de auxina este abolită și, de asemenea, expresia de OsBRI1 jos este reglementată în arf mutant (Sakamoto et al., 2013). S-a raportat că OsARF19 se leagă de promotorul OsBRI1 și reglează pozitiv expresia sa, care apoi activează semnalizarea BR (Zhang et al., 2015). BES1 poate lega la promotor de MICI AUXIN-UP ARN 15 (SAUR15) și să medieze BR devreme răspuns gene în Arabidopsis, iar acest lucru obligatorii ar putea fi îmbunătățită prin auxin tratament (Walcher și Nemhauser, 2012)., Luate împreună, auxina poate afecta, de asemenea, răspunsurile BR și creșterea și dezvoltarea plantelor reglementate de BR.în ultimele aproape patru decenii, studiile privind interacțiunile căii auxin-BR au atras tot mai mult interesul cercetătorilor. Aplicarea instrumentelor fiziologice, moleculare, genetice și biochimice ne-au aprofundat foarte mult înțelegerea acestei probleme., Pe baza studiilor anterioare, BR și auxin sunt implicate sinergic în multiple procese de dezvoltare a plantelor, inclusiv: alungirea hipocotilului, dezvoltarea fasciculelor vasculare, dezvoltarea rădăcinilor și tropismele etc. Interdependența și cooperarea dintre auxin și BR sunt complicate și implică numeroase procese la nivel molecular, prin împărtășirea acelorași gene țintă, reglându-se reciproc pe mai multe niveluri (Figura 1).
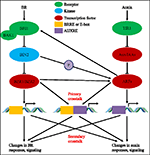
figura 1. Model de auxin-brassinosteroid (BR) crosstalk., În Arabidopsis, percepțiile semnalului BR și auxin sunt recunoscute de receptorii BRI1 și respectiv TIR1. BR se leagă de domeniul extracelular al BRI1 și de a promova interacționează cu co-receptorilor BAK1 pentru a forma o mai activă BR receptorilor complexe, care la rândul său duce la dephosphorylation și inactivarea BIN2. Inactivarea BIN2 duce la dephosphorylation de două BR omoloage factori de transcriere BZR1 și BZR2, care se mișcă în nucleu pentru a activa transcrierea unor gene care conțin BRRE sau E-box în regiunea promotorului., BIN2, de asemenea, pot fosforila ARF7 și ARF19 pentru a suprima interacțiunea lor cu AUX/IAAs și, astfel, a spori activitatea transcripțională pe gene tinta. TIR1 recepționează semnalizarea auxinei și interacționează cu proteinele aux / IAA ca co-receptor al auxinei. AUX/IAA apoi este degradat prin ubiquitin–proteazomal cale, iar auxina transcripțional reglementare auxin factorii de răspuns (Arf) sunt eliberate din AUX/IAA represiune și activează transcrierea unor gene cu auxina receptiv elemente (AUXRE) în activitatea de reglementare regiune., Unele ARF-uri se pot lega, de asemenea, de promotorul BRI1 și își reglează pozitiv expresia, care apoi activează semnalizarea BR. Crosstalk primar apare prin activarea genelor care conțin atât BRRE / e-box și AUXRE în regiunea lor promotor, permițând ambele căi de semnalizare pentru a reglementa direct transcriere. Crosstalk secundar apare prin exprimarea genelor care sunt fie auxin sau BR receptiv, dar activitățile care controlează expresia genelor care reglează răspunsul și semnalizarea altor hormoni.,
Fosforilare regulamentul joacă un rol crucial în BR cale de semnalizare, în special în procesul de percepție, BR este percepută prin BRI1-kinază și BAK1 kinaza co-receptori, și în cele din urmă controlează BR reglementate expresia genelor prin influențarea din aval factori de transcriere, cum ar fi BES1/BZR1 activități (A et al., 2005; Yin și colab., 2005; Sun et al., 2010; Tang și colab., 2011; Yu și colab., 2011). Cu toate acestea, reglementarea ubicuității pare esențială pentru semnalizarea auxinei., Odată auxina se leagă de TIR1 receptorilor, care acționează ca un E3 ubiquitin-ligaza, activat TIR1 E3-ligase ubiquitinates AUX/IAA proteine, duce la degradarea acestor repressors și de-a reprima ARF factori de transcriere, și în cele din urmă provoacă auxin reglementate expresia genelor schimbă modelul de creștere și răspunsuri (Gray et al., 1999, 2002; Hellmann și colab., 2003; Quint și colab., 2005). Deoarece s-a constatat că BIN2 kinaza, care este bine cunoscut funcționarea în BR semnalizare, poate fosforila și de a spori activitățile de Arf, cum ar fi ARF2 și ARF7 (Vert et al., 2008; Cho și colab.,, 2014), va fi interesant pentru a testa dacă kinaze cum ar fi BIN2, care sunt implicate în BR semnalizare, ar putea, de asemenea, interacționa cu alte auxin semnalizare componente, cum ar fi TIR1 receptorilor sau AUX/IAA repressors, și influența TIR1 E3-ligase activitate sau AUX/IAA proteine stabilitate. Pe de altă parte, rolul de ubiquitination în BR semnalizare, de asemenea, trebuie să fie abordate, mai ales dacă TIR1 E3-ligase putea interacționa direct cu BR de semnalizare componente și reglarea lor proteine stabilitate.,în plus, folosind auxin response DR5 și alți reporteri auxin, s-a observat că auxin reglează creșterea și dezvoltarea plantelor într-un mod dependent de țesut sau celular. Diferitele ieșiri transcripționale în funcție de contextul celular și de mediu (Clark et al., 2014; Etchells și colab., 2016; Lavy și colab., 2016)., Deși spațio-temporale BR semnalizare a fost dovedit pentru a controla creșterea rădăcinii prin acțiune antagonistă cu auxina (Chaiwanon și Wang, 2015), este încă necunoscut dacă tisulare sau celulare BR semnalizare, care ar putea fi vizualizate de către pBZR1:BZR1-YFP, este de asemenea important pentru a controla alte procese în afară de rădăcină de dezvoltare. În plus, generarea unui detaliate țesut sau pivniță harta de auxinelor și BR distribuții este posibil în prezent, fluorescence-activated cell sorting sau cu laser microdissection în combinație cu înaltă rezoluție expresia genelor analiză., Acest lucru va duce în cele din urmă la adresa dacă auxin crosstalks cu BR într-un țesut sau mod celular.
contribuții ale autorilor
toți autorii au fost implicați în redactarea acestui manuscris. Pentru mai multe informații despre ceea ce constituie autor, consultați ghidul nostru pentru autor.,
de Finanțare
Acest lucru este susținut prin subvenții de la National Science Foundation Naturale din China (Proiecte 31500229 și 31770305), Provincia Shandong Science Foundation Naturale Majore Bază Programul de Cercetare (2017C03), Shandong Provincial Fonduri pentru Distinși Tineri Cercetători (2014JQ201408).
Declarație privind conflictul de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Clouse, SD (2011)., Transducția semnalului Brassinosteroid: de la activarea kinazei receptorului la rețelele transcripționale care reglează dezvoltarea plantelor. Celula De Plante 23, 1219-1230. doi: 10.1105 / tpc.111.084475
PubMed Abstract | CrossRef Textul Complet | Google Scholar
Estelle, M. (2011). Auxin Signaling: de la sinteză la biologia sistemelor; o colecție de subiecte din perspectivele Cold Spring Harbor în biologie. Cold Spring Harbor, NY: rece primăvară Harbor laborator de presă.Google Scholar
Guilfoyle, TJ (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar