gena p53 a fost descoperită pentru prima dată în 1979. O proteină a fost identificată în virusul simian 40-celule de șoarece transformate (SV40) prin imunoprecipitare cu ser anti-T; această proteină a fost numită proteină p53 . În același an, Kress și alți oameni de știință au găsit o nouă clasă de proteine cu o masă moleculară cuprinsă între 50-60kda. Acest tip de proteină a fost apoi identificat ca p53 . Proteina p53 poate fi, de asemenea, identificată din diferite linii celulare transformate prin imunoprecipitare., Lane și Linzer au obținut, de asemenea, un rezultat similar în 1979. Alte dovezi pentru identificarea p53 este că p53 a fost exprimat în toate celulele de șoarece transformate testate; aceste teste includ sarcoame induse chimic, fibroblaste transformate și leucemii, în timp ce în celulele normale, p53 nu a fost exprimat. În plus, un nivel ridicat de p53 a fost detectat în majoritatea celulelor transformate, indiferent de modul în care celulele au fost transformate, fie spontan, fie non-spontan ., Acest lucru sa datorat în mare parte stabilității crescute a p53, cu toate acestea, în celulele carcinoa embrionare F9, a exprimat un nivel ridicat de p53, acest lucru sa datorat cantității de ARNm p53 tradus . după ce proteina p53 a fost descoperită în 1979, a devenit populară analiza acesteia. Cu toate acestea, la acea vreme, deoarece era o proteină recent descoperită și nu exista un nume anterior pentru aceasta, diferite instituții foloseau nume diferite și publicau lucrări cu nume diferite., Pentru a rezolva această problemă, în 1983, în timpul primului Atelier internațional p53 organizat în Oxted, Marea Britanie, oameni de știință din diferite grupuri de cercetare din diferite țări s-au reunit pentru a discuta o nomenclatură comună pentru această proteină recent descoperită. La această întâlnire, ” p53 ” a devenit numele său și a fost folosit de atunci. Sa crezut că motivul pentru care oamenii de știință au numit proteina p53 este că masa moleculară a acestei proteine este 53kDa care se bazează pe migrarea sa în gelul SDS. Ulterior, masa moleculară sa dovedit a fi greșită, iar masa moleculară corectă ar trebui să fie 43.,7kda deoarece p53 conține o regiune bogată în prolină, iar această regiune poate reduce migrarea p53 în gelul SDS. Dar numele ” p53 ” a rămas .
în anii 1980, se credea că proteina p53 este implicată în ciclul celular, precum și că joacă un rol în replicarea ADN-ului. Mai târziu, în 1982 până în 1994, oamenii au descoperit că unele oncoproteine virale au fost capabile să se lege de p53, formând un complex. În 1982, Sarnow și colab. a constatat că adenovirus E1b (58kDa) poate interacționa cu un 54kDa de proteine, care este prezent în SV40-a transformat celulele mouse-ul menționat mai sus., Conform rezultatelor specificităților imunologice ale anticorpilor T și hărților peptidice ale proteinei 54kda, această proteină 54kda este identificată ca p53 . În același an, oamenii de știință au descoperit că, dacă ar injecta anticorpul p53 în celulele elvețiene de șoarece 3T3, acesta ar inhiba celulele care intră în faza S a ciclului celular; în aceeași situație, anticorpul p53 nu a afectat SV40 sau sinteza ADN indusă de adenovirus .,
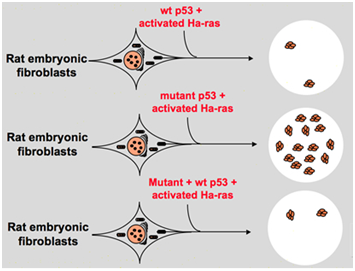
mai Târziu, în 1984, cercetătorii au analizat efectul p53 pe non transformat 3T3 fibroblaste; s-au analizat rata de sinteza a proteinei p53 la diferite momente de timp și a găsit că la sfârșitul G1 fază, rata de sinteza și nivelul proteinei p53 și legate de arnm crește. Acest rezultat sugerează că proteina p53 inhibă celulele care intră în faza de divizare din interfază . Maltzman W și colab. a făcut un alt experiment în același an. Ei au tratat celula de șoarece netransformată cu lumină UV și 4nqo carcinogen chimic UV-mimetic și au detectat un nivel ridicat de p53., Rezultatul a arătat că expresia ridicată a p53 nu este doar un simbol care indică ciclul celular, ci și o componentă mai importantă care este implicată în sinteza ADN și proliferarea celulelor . În 1987, când a studiat complexul de antigen t al virusului simian 40 și ADN polimeraza α, Gannon și alți oameni de știință au descoperit o schimbare similară a antigenului atunci când este legată de p53 și polimeraza α. Ei au descoperit, de asemenea, că la o anumită concentrație a celor trei componente, ele pot forma un complex trimeric special care include antigenul T, p53 și ADN polimeraza α., Deoarece antigenul T este implicat în replicarea ADN-ului viral și transformarea celulară, acest rezultat indică faptul că p53 joacă un rol în controlul ciclului celular și replicarea ADN-ului .după cum a arătat experimentul de mai sus, p53 are capacitatea de a imortaliza celulele. În 1984, Eliyahu D și colab. s-a constatat că p53 și produsul oncogene myc au împărtășit unele proprietăți similare. Ambele au capacitatea de a se lega de alte proteine și sunt implicate în ciclul celular și ambele se acumulează în nucleele celulelor transformate ., Bienz, Pennica și Oren analizat secvențele de aminoacizi a proteinei p53 și produsul dintre myc, și au descoperit că cele două proteine prezintă asemănări în structura moleculară și poziția speciale percepute reziduuri. Apoi, oamenii de știință au propus că p53 poate acționa ca o oncogenă. Pe baza acestei ipoteze, Eliyahu D și colab. am făcut niște experimente. Deoarece fibroblastele embrionare cu rată primară pot fi transformate prin implicarea atât a produsului myc, cât și a Ha-ras , celulele renale primare de șobolan pot fi, de asemenea, transformate prin cooperarea Ha-ras și adenovirus Regiunea timpurie 1A, Eliyahu D și colab., a decis să utilizeze acest tip de sistem de testare biologică pentru a identifica funcția oncogenă a p53. Ei au tratat celulele embrionare normale cu p53 și au activat Ha-ras. Rezultatul a arătat că celulele țintă se confruntă cu schimbări de morfologie și produc niveluri ridicate de p53, Eliyahu D și colab. crezut că transformarea fibroblastelor embrionare prin p53 și Ha-ras a explicat că gena p53 este o oncogenă ., În 1985, Jenkins a propus ca gena p53 să extindă durata de viață a celulelor, să sporească afectivitatea transformării prin rearanjarea secvenței sale de codificare care ar putea provoca producerea de proteine stabile .cu toate acestea, la sfârșitul anilor 1980, oamenii de știință au început să realizeze că p53 este o genă supresoare tumorală în locul unei oncogene. Ei au observat că p53 cu funcție normală nu poate fi detectată în multe dintre tumori și au constatat că pierderea expresiei și funcției genei p53 de tip sălbatic este necesară în timpul transformării celulare., Acestea cresc posibilitatea ca gena p53 de tip sălbatic poate inhiba progresia neoplazică . Apoi au formulat o altă ipoteză: gena clonă p53 utilizată în experimentele anterioare conține ocazional mutații negative dominante în domeniul extrem de conservat, ceea ce duce la rezultate opuse ale experimentului . În 1988, Ben și alți oameni de știință au detectat o cantitate mare de rearanjat p53 în murin eritroleucemia linii celulare–DP20-1 și CB3 care sunt derivate din splina de murine infectat cu Prietenul virus leucemie ., În 1989, Eliyahu, care a subliniat că p53 este o oncogenă și-a schimbat mintea și a presupus că gena p53 de tip sălbatic poate inhiba transformarea celulelor. Eliyahu și alți oameni de știință au studiat efectul proteinei p53 de tip sălbatic codificate de plasmide și p53 mutante asupra capacității de a obține rata primară de transformare a fibroblastelor embrionare prin diverse combinații oncogene in vitro. De exemplu mutant p53 plus ras și myc plus ras., Rezultatul a arătat că tipul sălbatic p53 duce la o reducere uriașă a focarelor transformate cauzate de mutantul p53 plus ras; mutantul p53 nu a arătat nicio inhibiție asupra focarelor transformate cauzate de myc plus ras, în timp ce transformarea mediată de myc plus ras este foarte sensibilă la expresia tipului sălbatic p53. Figura 1 arată acest experiment concis. Acesta a arătat că, în comparație cu p53 mutant, p53 de tip sălbatic prezintă un efect evident inhibitor asupra transformării celulare. Efectul este pozitiv legat de nivelul de Expresie al tipului sălbatic p53 și negativ legat de nivelul de Expresie al mutantului p53., Acest experiment a sugerat că tipul sălbatic p53 poate avea într-adevăr o funcție opusă în comparație cu p53 mutant și poate inhiba tumorigeneza . În prezent, p53 este recunoscut ca o genă supresoare tumorală. Se estimează că aproximativ jumătate din tumori sunt cauzate de p53. Este una dintre cele mai frecvent mutante gene la om și cea mai frecvent analizată genă din întreaga lume .
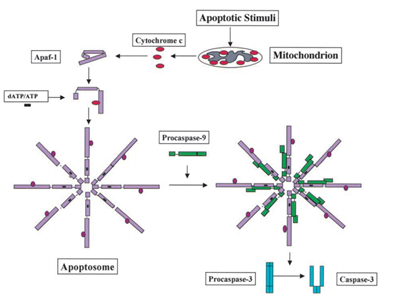
Mihara M și colab. oamenii de știință au descoperit că gena p53 de tip sălbatic poate fi translocată rapid pe suprafața mitocondrială a celulelor tumorale. În cadrul experimentului, au descoperit că unele proteine p53 de tip sălbatic induse de stres au capacitatea de a se transloca în mitocondriile timocitelor din celulele umane sau de șoarece după apoptoză din cauza deteriorării ADN și a hipoxiei. Apoi, aceste tip sălbatic proteine p53 induce permeabilizare de mitocondrii și poate provoca o serie de modificări care au loc în mitocondrii ca eliberarea citocromului c prin formarea de complexe cu Bcl2 și BclXL .,ca rezultat clinic bun, cu efect secundar redus, terapia genică este populară. Până la sfârșitul anului 2005, au existat 1020 de studii de terapie genică în baza de date a Journal of Gene Medicine. Printre aceste studii, 66% din terapiile genice au fost efectuate pe pacienți cu cancer, iar 58 de studii au folosit rAd-p53, un adenovirus recombinant care codifică gena p53 umană. În aprilie 2004, a fost lansată oficial o injecție recombinantă umană adenovirus-p53 (Gendicine). Gendicine este utilizat pentru a trata carcinomul cu celule scuamoase al capului și gâtului și a fost aprobat de Administrația de Stat pentru alimente și medicamente din China pe Octombrie. 16, 2003., A devenit primul produs de terapie genică din lume care a fost aprobat de guvernul chinez .gena p53 a fost descoperită pentru a regla metabolismul în 2005. Pentru a transfera de la faza G1 la S, celulele trebuie să aibă un suport suficient de materii prime pentru ADN, organele și sinteza proteinelor. Pentru a reglementa acest proces, sunt necesare câteva puncte de control. Unul dintre ele este punctul de control dependent de glucoză la G1/S. este reglat de protein kinaza activată de AMP (AMPK). Când glucoza este epuizată, AMPK poate fosforila proteina p53, care la rândul său induce arestarea celulelor și evită moartea celulelor., Celulele care se confruntă cu oprirea dependentă de p53 vor reintra în ciclul celular atunci când glucoza este suficientă .se știe că inactivarea p53 este necesară pentru formarea tumorilor. Bykov și colab. VJ și Snydel el și colab. subliniați că funcționarea necorespunzătoare a p53 poate duce la proliferarea unei tumori existente . Ventura și colegii săi au făcut câteva experimente pentru a testa această ipoteză. Ei au restabilit funcția p53 endogen în tumorile autohtone primare pentru a examina consecința reactivării p53., Rezultatul a arătat că reactivarea p53 a fost responsabilă pentru regresia tumorilor autohtone. Asta înseamnă că proteina p53 inactivată poate duce la dezvoltarea tumorii . Xue și alți oameni de știință au făcut, de asemenea, un experiment pentru a testa consecința reactivării p53 asupra tumorilor. Ei au folosit interferențe ARN reversibile (RNAi) pentru a regla expresia p53 endogen la soareci cu cancer la ficat. În experiment, doxiciclina (Dox) este utilizată pentru reactivarea p53, deoarece expresia p53 este suprimată total atunci când Dox lipsește și se restabilește rapid atunci când se adaugă Dox., Când a fost tratat cu Dox, miARN p53 a fost oprit, ceea ce, la rândul său, determină o expresie crescută a p53. Rezultatul a arătat că tumorile la șoarecii tratați cu Dox devin nedetectabile după 12 zile, în timp ce tumorile la șoarecii netratați au crescut rapid. Pentru a testa consecința reactivării tranzitorii a p53, au tratat șoarecii cu Dox timp de 4 zile și apoi s-au oprit. Rezultatul a arătat că chiar și un tratament de două zile poate provoca regresia tumorilor și 4 zile de tratament pot determina regresia completă a tumorilor., Ei au subliniat, de asemenea, că în timpul regresiei tumorale, p53 reactivat tranzitoriu poate declanșa senescența celulară, nu apoptoza. În același an, Hu a constatat că implantarea embrionară la șoarecii de sex feminin p53/ este reglată de factorul inhibitor al leucemiei (LIF). LIF este o citokină secretată și este importantă pentru implantarea blastocistului. Gena care codifică LIF este identificată ca gena țintă p53, iar situsul de legare p53 este localizat în intronul 1 atât la oameni, cât și la șoareci .