Úvod
na Rozdíl od zvířat, která se může pohybovat, aby se zabránilo nežádoucí okolí, přisedlé rostliny vykazují vysoce rozvinuté adaptaci na složité podmínky prostředí. K dosažení této hluboké přizpůsobivosti je nutná komunikace mezi buňkami. Komunikace mezi buňkami v rostlinách zahrnuje robustní zpracování intracelulární signalizace a složité mezibuněčné signalizační sítě., Až do teď tam jsou nejméně devět signalizace látek, jménem rostlinných hormonů, včetně auxin, brassinosteroid (BR), hodnotami, gibereliny (GA), ethylen, jasmonic acid (JA), strigolactone (SL), kyseliny abscisové (ABA), a kyselina salicylová (SA) objevil (Druege et al., 2016; Verma et al., 2016). Genetické a fyziologické studie odhalily kritické role a funkční mechanismy těchto výše uvedených hormonů v růstu a vývoji rostlin (Gray, 2004)., Na základě předchozí studie, auxin, BR, GA, SL, a hodnotami především funkci při normální růst a vývoj rostlin, zatímco ABA, etylén, JA a SA hrát důležitou roli v růstu rostlin, reakce na různé biotické a abiotické stresy (Pieterse et al., 2009; Santner et al., 2009; Denance et al., 2013). A také některé z těchto hormonů mají dvojí roli, například ABA také hraje důležitou roli ve vývoji semen a klidu (Seo a Koshiba, 2002)., Ačkoli každý hormon hraje v určitých aspektech převládající roli, mnoho hormonů překrývalo aktivity a interakce různých hormonů řídí mnoho vývojových aspektů a růst v reakci na endogenní vývojové a exogenní podněty.
Auxin a BR jsou dvě hlavní třídy hormonů podporujících růst., BR, skupina rostlin-specifické steroidní hormony, které by mohly komunikovat s ostatními fytohormonů jako je auxin, hodnotami, ethylen, GA, JA a SA a regulovat širokou škálu rostlinných růstových a vývojových procesů, včetně klíčení semen, buněčné prodloužení, cévní diferenciace, průduchy vznik a pohyb, květu a mužské plodnosti (Saini et al., 2015). Je zajímavé, že každý z těchto procesů je také řízen auxinu, což naznačuje, že tyto dva hormony souhra kontrolovat vývoj rostlin., V této recenzi, jsme se nastínit přenosu signálu auxinu a BR na základě nedávného pokroku a přezkoumání přeslechy mezi auxinu a BR zprostředkovaný růst a vývoj rostlin.
Auxinová signalizační dráha
Auxin byl poprvé rozpoznán jako rostlinný hormon kvůli své roli v tropismu rostlin na gravitaci nebo světelné podněty., Později auxin byl chemicky identifikován jako indol-3-octové kyseliny a prokázáno, že hrát zásadní roli v nepřeberné množství rostlinných vývojových a fyziologických procesů, včetně embryogeneze, organogeneze, cévní diferenciace, kořen a střílet rozvoj, tropic růst a vývoj plodu (Estelle, 2011).
pomocí genetické analýzy v Arabidopsis byl molekulární mechanismus, který je základem auxinové signální transdukce, dobře prozkoumán. Transportní inhibitor RESPONSE1 (TIR1) byl první identifikovaný jaderný receptor auxinu (Ruegger et al., 1998; Dharmasiri et al., 2005)., TIR1 kóduje jaderný protein patřící k proteinu F-box jako podjednotku proteinového komplexu SCF E3 ubiquitin ligázy (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005), kromě TIR1, tam jsou další tři F-box proteinů a to AUXIN SIGNALING F-BOX Proteinů (AFBs), které ukazují, auxin-binding aktivitu a zprostředkovat auxin signalizace u Arabidopsis (Badescu a Napier, 2006). Receptor TIR1 může interagovat se skupinou proteinů AUX/IAA (auxin/indol-3-octová kyselina) (Dharmasiri et al., 2003)., Proteiny AUX/IAA jsou negativními regulátory auxinové signalizace a v genomu Arabidopsis je zakódováno 29 členů AUX/IAA. AUX/IAA proteiny může interagovat s třídou transkripční regulátory, auxin response factors (ARF), zprostředkující transkripční reakce k auxinu. Pod vysokým auxin úrovni, AUX/IAA proteiny interagují s TIR1 jako coreceptor auxinu, a může být ubiquitní do SCFtir1 složité, a tak bude degradován prostřednictvím ubiquitin–proteazomové dráhy (Gray et al., 2001; Lanza et al., 2012)., Po zničení AUX/IAA repressors, auxin transkripční regulátory ARFs, které zahrnují 23 členství může být propuštěn z AUX/IAA represe, a tak zprostředkovat auxin reakce aktivace nebo represe cílových genů (ním matka mluvila, říkal, a Hagen, 2007). Různé sady F-box bílkovin a AUX/IAA nebo ARFs odvodit složitosti během auxin signální transdukce (Goh et al., 2012; Guilfoyle, 2015; Salehin et al., 2015).,
koordinované působení transkripčních represorů Aux / IAA a transkripčních faktorů ARF vytváří komplexní genově regulační sítě, které byly také hlášeny ve Physcomitrelle (Lavy et al., 2016). Nedávno bylo zjištěno, že podjednotka SCF CULLIN1 (CUL1) interaguje s TIR1 a reguluje tak stabilitu substrátů TIR1 a signalizaci auxinu (Wang et al., 2016). Interakce mezi TIR1 a Aux/IAA je také ovlivněn prostorové konformace Aux/Iaa, řízen cyclophilin izomeráza LRT2 v rýži (Jing et al., 2015)., HEAT SHOCK FACTOR 90 (HSP90) a co-garde SGT1, respektive, spolupracuje s TIR1 a tím reguluje TIR1 stabilitu, která ovlivňuje interakce mezi TIR1 a Aux/IAA a auxin signalizace (Wang et al., 2016).
Kromě TIR1-závislé kanonické auxin-signální dráhy, auxin bylo nedávno oznámeno, vyvolat pestrou škálu vývojových odpovědi prostřednictvím non-kanonický auxin-signalizační mechanismus., V této non-kanonický auxin snímací proces, ARF3/ETTIN kontroly genové exprese prostřednictvím interakce s procesu-specifické transkripční faktory, což velmi obohacuje auxin-zprostředkované rostlin vývojové rozmanitosti (Simonini et al., 2016, 2017).
br signalizační dráha
BRASSINOSTEROID byl poprvé objeven v pylu pro svou schopnost podporovat prodloužení buněk. Později bylo zjištěno, že BR hraje roli v široké škále aspektů růstu rostlin a může reagovat na biotické a abiotické stresy., V dnešní době byla br signální transdukční dráha do značné míry objasněna kombinací různých metod, včetně molekulární genetiky, biochemie, proteomiky a genomiky atd. Buňka-povrch kinázy BRASSINOSTEROID INSENSITIVE1 (BRI1) byl identifikován jako receptor pro BR, která se může vázat na extracelulární domény BRI1 a aktivovat jeho kinázy a tím přepnout na signální kaskády pro regulaci transkripce (Li a Chory, 1997; Wang et al., 2001; Kinoshita et al., 2005; Kim a Wang, 2010; Clouse, 2011; Hothorn et al., 2011; ona et al., 2011; oh et al., 2012)., Při vnímání BR, BRI1 interaguje s co-receptoru BRI1-SPOJENÉ KINÁZY 1 (BAK1) a jeho homolog SOMATICKÉ EMBRYOGENESISRECEPTOR Kináz (SERKs) tvoří více aktivní BR receptorového komplexu (Li et al., 2002; Nam and Li, 2002; Wang et al., 2005; Tang et al., 2008; Gou et al., 2012). Aktivuje BRI1 fosforyluje dva substráty plazmatické membrány-kotví receptor-jako cytoplazmatické kináz: BRASSINOSTEROID-SIGNALIZACE KINASES1 (BSK1) a KONSTITUTIVNÍ DIFERENCIÁLNÍ GROWTH1 (CDG1) (Tang et al., 2008; Kim et al.,, 2011), který následně fosforyluje a PP1-typ fosfatázy jménem BRI1-SUPPRESSOR1 (BSU1) pro aktivaci BSU1, což vede k BSU1 dephosphorylation a inaktivace na GSK3-like kinázy BRASSINOSTEROID INSENSITIVE2 (PŘIHRÁDKA 2). Kinázová aktivita BIN2 je také inhibována Histon DEACETYLÁZOU HDA6, která interaguje a deacetyluje na K189 BIN2., Když BR úrovně jsou nízké, BRI1 je v klidu vzhledem k jeho negativním regulátorem, BRI1 KINÁZY 1 (BKI1) a protein fosfatázy 2A (pb 2a), zatímco PŘIHRÁDKA 2 fosforylovat dva BR homologické transkripční faktory, BRASSINAZOLE RESISTANT1 (BZR1) a BZR2 (také jmenoval BES1 pro BRI1-EMS-SUPRESOROVÝCH 1) (Mu et al., 2002; Wang et al., 2002; Yin et al., 2002; Mora-Garcia et al., 2004; Kim et al., 2009, 2011; Kim a Wang, 2010). Když BR úrovně jsou vysoké, PŘIHRÁDKA 2 je inaktivovaná, a BZR1 a BZR2 jsou defosforylována tím, pb 2a, a přesuňte do jádra změnit výraz tisíce BR reakci genů (He et al.,, 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011).
synergie mezi signalizací BR a auxinu
signální dráhy auxinu a BR hrají různé role, ale také vykazovaly synergické a vzájemně závislé interakce v široké škále vývojových procesů. Například signály auxinu i BR mohou podporovat expanzi buněk a mohou synergicky interagovat na podporu prodloužení hypocotylů (Nemhauser et al., 2004)., Reakce jedné ze dvou drah v podpoře hypokotylu prodloužení vyžaduje funkci druhého a vzájemnou závislost mezi BR a auxin drah (Nemhauser et al., 2004). Auxin zvýšil hypokotylu délka wild-type rostliny, ale ne v BR-mutant necitlivý bri1-116, a to auxin-necitlivé fenotyp bri1-116 byla potlačena dominantní gain-of-function mutant bzr1-1D, což naznačuje, BR nebo aktivní BZR1 je nutné pro auxin propagace hypokotylu prodloužení., Bylo zjištěno, že BR signalizace konverguje s TLUMIČEM PHYTOCHROME B4-3 (SOB3) pro kontrolu buněčné prodloužení a hypokotylu růst prostřednictvím nařízení auxinu indukované MALÉ AUXIN SE RNA19 (SAUR19) výraz (Favero et al., 2017). Na druhou stranu, auxin regulované transkripční faktor MALÝ ORGÁN VELIKOSTI 1 (SMOS1) nedávno bylo zjištěno, že kontrolní buněčné expanze prostřednictvím přímé interakce s SMOS2/DLT, člen GRAS rodiny transkripční ko-regulátorů, která hraje pozitivní roli v BR signalizace v rýži (Kim et al., 2009; Tong et al., 2012; Hirano et al., 2017)., Auxin související mutanti jako iaa3 a arf6/arf8 byly méně citlivé na BR než byl wild-type pro prodlužování hypokotylu, a zrušil přecitlivělost bzr1-1D k auxinu, což naznačuje, že BR a BZR1 propagace hypokotylu prodloužení vyžaduje ARF6/8. Genomu-široký ChIP-Seq analýza ukázala, že ARF6 sdílí obrovské množství genových cílů (kolem 50%) s BZR1 a světlo/teplota-regulovaný transkripční faktor PIF4 tím, CHIP-Seq analýzy (Oh et al., 2014)., BZR1 a PIF4 interagují s ARF6 a aktivují sdílené cílové geny vazbou na sdílené cílové geny kooperativně během prodloužení hypokotylů (Oh et al., 2014) a mnoho z těchto překrývajících se cílových genů kóduje proteiny buněčné stěny zapojené do buněčné expanze.
Brassinosteroid a auxin také hrají důležitou roli v udržování kořenový apikální meristém (RAM) (Durbak et al., 2012). RAM se skládá z malé skupiny zřídka dělících buněk známých jako klidové centrum (QC), obklopené kmenovými buňkami, které vedou k různým typům tootových tkání., Udržování populace kořenových kmenových buněk je regulováno HOMEOBOXEM souvisejícím s WUSCHELEM 5 (Wox5) (Sarkar et al., 2007). Wox5 je omezen na QC auxinovou signalizací a usnadňuje správnou expresi PLT genů (Aida et al., 2004; Ding a Friml, 2010). Mutace genu br receptoru BRASSINOSTEROID necitlivý 1 (BRI1) vést k aberantní progrese buněčného cyklu v paměti RAM a způsobit menší beranů (Gonzalez-Garcia et al., 2011; Hacham et al., 2011). Auxin je známo, že stimuluje biosyntézu BR (Chung et al.,, 2011), ale aktivita BR neovlivňuje expresi PIN genů (Hacham et al., 2011). Fenotypy kořenového hrotu mutantů BR nevykazují totéž jako auxinové mutanty (Gonzalez-Garcia et al., 2011), což naznačuje, že BR působí na RAM nezávisle na auxinu.
Brassinosteroidní a auxinové signály jsou také synergicky vyžadovány při tvorbě radiálních vzorců cévních svazků (Ibanes et al., 2009)., U kombinací matematického modelování a biologické experimenty, auxin maxima, zřízený asymetrické polar auxin transport, ale nejsou změny na auxin úrovně je důležité pro umístění cévních svazků. Ukázalo se, že signál BR slouží jako podpůrný signál pro počet buněk v provaskulárním kruhu, které jsou v souladu s auxin maxima. Tedy vytvoření pravidelné uspořádání cévních svazků v natáčení je v rámci koordinovaného působení těchto dvou rostlinných hormonů (Ibanes et al., 2009)., Oba signály se také podílejí na vývoji kořenů rostlin a interakce BR a auxinu je zprostředkována BREVIS RADIX (BRX) během tohoto procesu. BRX je důležitý pro biosyntézu br a BR exogenní aplikace, která může zachránit mutantní defekty brx. Kromě toho, auxin-citlivý genové exprese je celosvětově postižené v brx mutant, a expresi BRX je silně indukována auxinu a potlačena BR, což znamená, BR biosyntézu auxinu a signalizace jsou připojeny přes smyčku zahrnující BRX během kořenový rozvoj (Mouchel et al., 2006).,
brassinosteroidy a auxin také hrají synergické role během vývoje postranních kořenů. BRs fungují hlavně při iniciaci laterálního kořene primordia, zatímco auxin je vyžadován jak pro iniciační, tak pro fáze vzniku laterální tvorby kořenů (Casimiro et al., 2001; Bhalerao et al., 2002; Benkova et al., 2003; Bao et al., 2004). Během těchto procesů BRs zvyšují iniciaci LRP podporou transportu auxinu acropetalu v kořeni, ale neovlivňují hladinu endogenního IAA (Bao et al., 2004)., Všechny tyto zprávy naznačují, že přeslechy mezi BR a auxin hraje důležitou roli v regulaci růstu rostlin a vývoje.
BR Auxin Reguluje Signalizaci
Kromě vzájemné závislosti a spolupráce auxinu a BR signálů během vývoje rostlin, BR by mohl zprostředkovat auxin signál dráhy na více úrovních. BZR1 interaguje s ARF proteiny se přímo zaměřují více auxinu signalizačními prvky a geny zapojené do metabolismu auxinu, jako jsou doprava a signalizace, včetně AUX/IAA, Kolíky, TIR1, a ARFs, atd. (Sun et al., 2010)., Bylo zjištěno, že Aux/IAA proteiny jsou zapojeny v BR odpovědi a iaa7/axr2-1 a iaa17/axr3-3 mutanty vykazovaly aberantní BR citlivost a nenormální BR-indukované genové exprese v orgánu způsobem závislým (Nakamura et al., 2006). Exogenní léčba brassinolidem (BL) by mohla vyvolat expresi genů reagujících na auxin, jako jsou iaa5, IAA19, IAA17 atd. a výraz z výše uvedených genů je regulována dolů v BR biosyntetických mutant de-etiolated2 (det2), což naznačuje, že funkční BR biosyntéza je částečně nutné pro auxin-dependentní genové exprese (Nakamura et al.,, 2003; Kim et al., 2006). Kromě toho BR také ovlivňuje tok auxinu regulací výrazu vývozců auxinu, jako jsou PIN4 a PIN7 (Nakamura et al., 2004). Během závodu gravitropism odpovědi, BRs mohl zvýšit polární akumulace auxinu vývozce PIN2 v kořenovém meristému, zóny a tím ovlivňují redistribuci auxinu z kořene tip k prodloužení zón a výsledek na rozdíl IAA úrovně v obou horní a dolní strany kořeny, přimět rostliny gravitropism., Během tohoto procesu hraje BR aktivovaný ROP2 důležitou roli při modulaci funkční lokalizace PIN2 regulací sestavy / opětovné montáže F-aktinů (Li et al., 2005). Další studie ukázaly, že snížené vnímání BL a/nebo koncentrace by mohly indukovat cyp79b2, gen kódující enzym konvertující tryptofan na indol-3-acetaldoxim, a tak ovlivnit distribuci (Kim et al., 2007).
kromě toho bylo zjištěno, že signál BR může regulovat signalizační výstup auxinu svým negativním regulátorem GSK3 kináza BIN2., Faktor odpovědi auxinu ARF2 byl identifikován jako protein interagující s BIN2 v kvasinkové dvou-hybridní obrazovce a test kinázy ukázal, že BIN2 může fosforylovat ARF2. Fosforylace ARF2 vede ke ztrátě vazebné schopnosti DNA a represivní aktivity cílových genů (Vert et al., 2008). ARF2 je BZR1 cílový gen a jeho exprese je redukována léčbou BR (Sun et al., 2010)., Navíc, PŘIHRÁDKA 2 může fosforylovat ARF7 a ARF19 potlačit jejich interakce s AUX/Iaa a tím zvyšují transkripční aktivitu cílových genů BOČNÍ VARHANY HRANICE-DOMAIN16 (LBD16) a LBD29 regulovat boční kořen organogeneze (Cho et al., 2014). BR však hraje během tohoto procesu malou roli a BIN2 je pod kontrolou modulu inhibičního faktoru diferenciace TRACHEÁRNÍHO prvku (TDIF) – tdif RECEPTOR (TDR) (Cho et al., 2014). Společně může BR regulovat auxinové reponse ovlivněním různých signalizačních složek auxinu.,
Auxin reguluje signalizaci BR
na druhé straně auxin může v určitých aspektech také regulovat signální dráhu BR. Exprese DWARF4, rozhodující hydroxylázy pro biosyntézu BR pro kontrolu endogenní hladiny BR, je závislá na auxinu. Auxin léčba by mohla výrazně stimulují expresi DWARF4 a auxin může inhibovat vazbu BZR1 se pořadatelem DWARF4. Indukce DWARF4 auxinem vyžaduje auxinovou signalizační dráhu, ale ne signální dráhu BR (Chung et al., 2011; Yoshimitsu et al., 2011)., CPD katalyzující C-3 oxidace BR byla aktivována BRX, domnělý transkripční faktor působící po proudu auxinové signalizace (Mouchel et al., 2006). Další studie v rýži naznačuje, že exogenní auxin může zvýšit hladiny exprese transkripce genu br receptoru OsBRI1, což naznačuje, že auxin zvyšuje signalizaci BR regulací br receptorů (Sakamoto et al., 2013). Kromě toho má promotor OsBRI1 motiv upstream auxin-response element (AuxRE), který je zaměřen na transkripční faktory ARF., Mutantní studie navíc naznačují, že po mutaci AuxRE je zrušena indukce exprese OsBRI1 auxinem a také exprese OsBRI1 je u ARF mutantu (Sakamoto et al., 2013). Bylo hlášeno, že OsARF19 se váže na promotor OsBRI1 a pozitivně reguluje jeho výraz, který pak aktivuje signalizaci BR (Zhang et al., 2015). BES1 se může vázat na promotor MALÝCH AUXIN-UP RNA 15 (SAUR15) a zprostředkovat BR včasné reakce genů v Arabidopsis, a tato vazba by mohla být posílena tím, že auxin léčby (Walcher a Nemhauser, 2012)., Společně auxin může ovlivnit i růst a vývoj rostlin.
závěrečné poznámky a budoucí perspektiva
během posledních téměř čtyř desetiletí přitahovaly studie interakcí auxin-BR dráhy stále větší zájem vědců. Přístroj fyziologických, molekulárních, genetických a biochemických nástrojů výrazně prohloubil naše chápání tohoto problému., Na základě předchozích studií jsou BR a auxin synergicky zapojeny do více vývojových procesů rostlin, včetně: hypocotyl elongace, vývoje cévních svazků, vývoje kořenů a tropismů atd. Provázanost a spolupráce auxinu a BR jsou komplikované a zahrnují četné procesy na molekulární úrovni, sdílením stejných cílových genů, regulaci vzájemně na více úrovních (viz Obrázek 1).
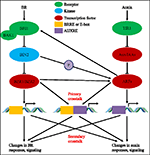
obrázek 1. Model přeslechů auxin-brassinosteroid (BR)., V Arabidopsis jsou vnímání signálu BR a auxinu rozpoznávány receptory BRI1 a TIR1. BR se váže na extracelulární doménu BRI1 a podporovat ji v interakci s co-receptor BAK1 tvořit více aktivní BR receptor komplex, který v zase vede k dephosphorylation a inaktivace PŘIHRÁDKA 2. Inaktivaci PŘIHRÁDKA 2 vést k dephosphorylation dvou BR homologické transkripční faktory BZR1 a BZR2, které se pohybují v jádře k aktivaci transkripce genů obsahující BRRE nebo E-box v jejich promotorové oblasti., BIN2 také může fosforylovat ARF7 a ARF19 potlačit jejich interakci s AUX / IAAs a tím zvýšit transkripční aktivitu na jejich cílových genech. TIR1 přijímá auxinovou signalizaci a interaguje s proteiny AUX/IAA jako co-receptor auxinu. AUX/IAA pak je degradován prostřednictvím ubiquitin–proteazomové dráhy, a auxin transkripční regulátory auxin response factors (ARFs) jsou uvolňovány z AUX/IAA represe a aktivaci transkripce genů s auxin responsive elements (AUXRE) v jejich regulační oblasti., Některé ARFs může se také váže na promotor BRI1 a pozitivně reguluje jeho výraz, který pak aktivuje BR signalizace. Primární přeslechy dochází aktivací genů, které obsahují jak BRRE/E-box a AUXRE v jejich promotorové oblasti, což obě signální dráhy přímo regulují transkripci. Sekundární přeslechy dochází prostřednictvím exprese genů, které jsou buď auxin nebo BR citlivý, ale činnosti, které kontrolují expresi genů, které regulují reakci a signalizace jiných hormonů.,
Fosforylace nařízení hraje klíčovou roli v BR signální dráhy, zejména během proces vnímání, BR je vnímán prostřednictvím BRI1 kinázy receptoru a BAK1 kinázy co-receptory, a nakonec kontrol BR regulované genové exprese prostřednictvím ovlivňování následné transkripční faktory, jako jsou BES1/BZR1 činnosti (He et al., 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011). Zdá se však, že regulace ubiquitinace je nezbytná pro signalizaci auxinu., Jednou auxin se váže na receptor TIR1, který působí jako E3 ubiquitin-ligázy, aktivní TIR1 E3-ligázy ubiquitinates AUX/IAA proteiny, vede k degradaci těchto repressors a de-potlačuje transkripční faktory ARF, a nakonec způsobí, že auxin regulované genové exprese vzor změny a růstové odezvy (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005). Od té doby bylo zjištěno, že PŘIHRÁDKA 2 kinázy, která je dobře známá fungování v BR signalizace, může fosforylovat a zlepšit činnosti ARFs jako ARF2 a ARF7 (Vert et al., 2008; Cho et al.,, 2014), bude zajímavé vyzkoušet, jestli kináz jako PŘIHRÁDKA 2, které jsou zapojeny v BR signalizace, mohl také komunikovat s ostatními auxin signaling komponenty jako receptor TIR1 nebo AUX/IAA repressors, a vliv TIR1 E3-ligázy činnosti nebo AUX/IAA proteinů stabilities. Na druhou stranu je třeba řešit také roli ubiquitinace v BR signalizaci, zejména pokud by TIR1 E3-ligáza mohla přímo interagovat s br signalizačními složkami a regulovat jejich proteinové stability.,
kromě toho, pomocí auxin response DR5 a další auxin novinářům, bylo zjištěno, že auxin reguluje růst a vývoj rostlin v tkáni nebo buněčné způsobem závislým. Různé transkripční výstupy v závislosti na buněčném a environmentálním kontextu (Clark et al., 2014; Etchells et al., 2016; Lavy et al., 2016)., Když prostoročasové BR signalizace bylo prokázáno, že ovládat růst kořenů prostřednictvím antagonistické působení s auxinu (Chaiwanon a Wang, 2015), to je stále neznámý, a pokud tkáňové nebo buněčné BR signalizace, které by mohly být vizualizovány pBZR1:BZR1-YFP, je také důležité řídit další procesy kromě kořenový rozvoj. Dále, generace podrobný tkáně nebo ve sklepě mapa auxinu a BR distribuce je v současné době možné pomocí fluorescence-activated cell sorting nebo laser microdissection v kombinaci s vysokým rozlišením genové exprese analýzy., To nakonec vede k řešení, pokud auxin crosstalks s BR v tkáni nebo buněčným způsobem.
autorské příspěvky
všichni autoři byli zapojeni do psaní tohoto recenzního rukopisu. Další informace o tom, co představuje autorství, naleznete v našich autorských pokynech.,
finanční Prostředky
Tato práce je podporována granty z Národní Přírodní Science Foundation Číny (Projekty 31500229 a 31770305), Provincie Šan-tung Přírodní Science Foundation Hlavní Základní Výzkum Program (2017C03), Shandong Provinční finanční Prostředky za vynikající Mladých Vědců (2014JQ201408).
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Clouse, S. D. (2011)., Brassinosteroid signální transdukce: od aktivace receptor kinázy po transkripční sítě regulující vývoj rostlin. Rostlinná Buňka 23, 1219-1230. doi: 10.1105 / tpc.111.084475
PubMed Abstract / CrossRef Full Text / Google Scholar
Estelle, m. (2011). Auxinová signalizace: od syntézy po systémovou biologii;sbírka předmětů z perspektivy studeného jara v biologii. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Google Scholar
Guilfoyle, T. J. (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar