Úvod
Prasátko úmrtnost je jedním z hlavních výběr vlastnosti v chovu prasat a je ovlivněna prasnice, selata, a na životní prostředí. Proto je úmrtnost prasat komplexním fenotypem a závisí na schopnosti prasnice vychovávat své potomky, ale je také funkcí porodní hmotnosti, řízení a výběru (Knol et al., 2002)., K úmrtnosti prasat však přispívají také monogenní recesivní vady, i když v minulosti bylo hlášeno pouze několik příkladů (Murgiano et al., 2012; Matika et al., 2019). Dokonce i v těch případech, kdy je účinek mutace závažný, je efektivní výběr proti takové mutaci omezen nízkou frekvencí. V mnoha závažných vad, zygoty zemřít velmi brzy v těhotenství, beze stopy, jiné než absence homozygotů v populaci (Derks et al., 2019).,
účinky příbuzenské Plemenitby v komerční populace prasat jsou obvykle uchovávány v šachu tím, že selektivní chov pro snížení mortality u selat zlepšením i mateřství schopnosti a prasátko životaschopnosti (Olijslagers, 2018). Nicméně, varianty hlubších recesivních monogenních vad, nejsou dobře zachyceny v rámci plemenné hodnoty, a potenciálně drift na vyšších frekvencích v důsledku intenzivní selekce (Georges et al., 2019). Kromě toho mohou být tyto varianty také udržovány v důsledku vyvažování výběru pro korelovaný pozitivní účinek v heterozygotním stavu (Derks et al., 2018).,
recesivní vady jen nepatrně přispívají k celkové úmrtnosti prasat (Alonso-Spilsbury et al., 2007). Nicméně varianty ovlivňující úmrtnost prasat mají velký význam, protože tyto varianty přímo ovlivňují produkci a dobré životní podmínky zvířat (Baxter et al., 2013; Rutherford et al., 2013). Při řízení populace zvířat je však nízkofrekvenční výskyt defektů obvykle špatně zdokumentován (často se používají velmi obecné termíny) a syndromy jsou často rozpoznávány pouze tehdy, když dosáhnou vysoké frekvence., To je zvláště důležité pro syndromy, které nevedou k velmi odlišným fenotypům. Proto i v komerčních chovných populacích lze provést malé sledování specifických syndromů a efektivní výběr proti specifickým nízkofrekvenčním syndromům proto vyžaduje nové přístupy.
V této práci jsme popsali objev vysoce oslabující syndrom v komerční populace prasat prostřednictvím průzkumu založeny na kombinaci medium-density SNP arrays a whole-genome sequencing (WGS)., Průzkum vedl k identifikaci delece 16-bp frameshift v genu SPTBN4, s předpokládanými jasnými fenotypovými důsledky u homozygotů. Nosná frekvence je ve sledované populaci asi 9%, což postihuje přibližně 0,81% populace. Frekvence byla dostatečně nízká, aby bylo známo, že mají genetický základ, a dokonce i efektivně, že nerozpoznané jako specifický syndrom. Po provedení průzkumu byla identifikována jedna těhotná prasnice, která byla vypálena nosným kancem., Postižená selata trpí myopatií a jsou schopni chůze, což má obvykle za následek smrt během několika hodin po porodu, plně v souladu s předpokládanými patologie ve srovnání s podobnou lidskou a myší případech.
Materiál a Metody
Zvířata, Genotypů, a Pre-Zpracování
dataset se skládá z 31,839 zvířata ze syntetického kance souladu s velké bílé pozadí. Linka je udržována a chována v Topigs norsvin Nucleus farms, především výběrem výrobních a zdravotních vlastností., Zvířata byla genotypizována na vlastním čipu Illumina GeneSeek 50K SNP (Lincoln, NE, USA). Zvířata s četností chybějících genotypů > 0.15 byla odstraněna. Jsme zlikvidovat značky, které nesplňují následující kritéria filtrování: minimální sazba 0,85, menší frekvenci alel > 0.01, a Hardy-Weinbergova poměru exaktní test p-hodnota je nižší než P < 10-12. Kromě toho byly značky s neznámým umístěním na sestavení genomu Sscrofa11.1 vyřazeny a po filtrování zůstalo 41 573 markerů., Všechny kroky byly provedeny v Plink v1.90b3.30 (Purcell et al., 2007).
Haplotyp Fázování a Identifikace SSC6 Haplotyp
provedli Jsme haplotyp fázování a imputace chybějících míst v Beagle5.0 s parametrem pro efektivní velikost populace nastavena na 100, ostatní nastavení byla výchozí (Browning et al., 2018). Očekávané homozygoty byly odhadnuty na základě haplotypové frekvence pomocí principu Hardy-Weinberg. Pro testování počtu pozorovaných homozygotů s počtem očekávaných homozygotů byl použit přesný binomický test., Haplotyp byl považován za významně vyčerpaný, pokud P < 5 × 10-3.
Fenotypové Účinky Spojené S SSC6 Haplotyp
zkoumali Jsme SSC6 haplotyp pro záznamy na celkový počet narozených, počet mrtvě narozených, mumifikovaná selata, porodem přežití, a kojení přežití (přežití až o 21 dní věku) celkem 9,666 vrhy. Tyto fenotypy jsme uvedli pro všechny identifikované vrhy CxC a cxn. Použili jsme welchův t-test k posouzení, zda se fenotypy z vrhu CxC výrazně liší od vrhu cxn. Hodnota p < 0.,05 byl považován za významný.
analýza sekvenování celého genomu a identifikace kandidátní varianty
dataset se skládá ze 71 jedinců sekvenovaných celým genomem ze studované populace. Všech 71 vzorků, které byly také přítomny v našem souboru dat 31,839 zvířat s genotypem na 50K. 71 vzorků, které mají celkový objem 1.93 Tbp (tera párů bází) od 14.16 miliardy 150-bp párových-koncových čte (Tabulka S3). Vzorky byly sekvenovány na Illumina HiSeq 2000. Sekvence jsme zarovnali na sestavení genomu Sscrofa11.1 pomocí verze BWA-MEM 0.7.,15 (Li and Durbin, 2009) s průměrnou mappability z 98.9% a vzorek pokrytí v rozmezí od 8.8 do 14.8 X (10.9 X průměr). Samblaster byl použit k odstranění duplikátů PCR (Faust and Hall, 2014). Samtools byl použit pro třídění, sloučení a index bam soubory (Li et al., 2009). Mapování a statistiky kvality byly generovány pomocí Qualimap (Okonechnikov et al., 2016). Varianta volání byla provedena s Freebayes v1.1.0 s následující nastavení: –min-base-kvalitní 10 –min-alternativní-zlomek 0.2 –haplotyp-délka 0 –min-alternativní-count 2 (Posádka a Marth, 2012)., Varianty se skóre kvality Phred < 20 byly vyřazeny (Li et al., 2009). Varianty byly anotovány pomocí predikátoru ensembl variant effect (VEP, release 96) (Mclaren et al., 2016). Dopad variant missense byl předpovězen pomocí třídění netolerantní od tolerantní (SIFT) (Kumar et al., 2009). LD analýza byla provedena za použití Plink v1. 90b3.30 (Purcell et al., 2007) s následujícími nastaveními –chr-set 18, –r2, ld-window-r2 0.8.
zarovnání proteinů Sptbn4
zarovnání proteinů mezi divokým typem a mutovaným proteinem bylo provedeno pomocí ClustalO (Madeira et al.,, 2019) a vizualizován pomocí ESPript 3 (Robert A Gouet, 2014). Další vizualizace a validace byla provedena pomocí jbrowse genome viewer verze 1.12.1 (Skinner et al., 2009).
Ověření Příčinné 16 bp SPTBN4 Smazání
PCR byla provedena pomocí 60 ng genomové DNA, 0,4 µm každého primeru, 1,8 mM MgCl2 a 25 jednotek/ml OneTaq® DNA Polymerase (OneTaq® 2X Master Mix s Standardní Pufr New England Biolabs) výrobce PCR buffer, které jsou v konečném objemu 12 µl., Počáteční denaturace 1 min při 95°C, následovalo 35 cyklů 95°C po dobu 30 s, 55°C po 45 s, 72°C 90 s, následuje 5 min. prodloužení 72°C. PCR primery pro SPTBN4 jsou TCAAGGGTGCAGGCTCTTTC dopředu a GGTAGGAAGCTCGAAGTGGG zvrátit. Forward primer byl barvivo označené buď 6-FAM k výrobě fluorescenčně značeného PCR produktu detekovatelné na ABI 3730 sekvenceru DNA (aplikované biosystémy). Velikosti fragmentů byly stanoveny pomocí softwaru GeneMapper 5 od společnosti ABI.,
Histopatologické Vyšetření
Dva postižených selat méně než 1 týden staré byly odeslat na oddělení patologie Královské Zdraví Zvířat (Deventer) pro vyšetření. Makroskopicky byla všechna pozorování v normálních mezích. Pro rutinní barvení H&e a barvení Ptah byl odebrán vzorek kosterního svalu přední části, hřbetního svalu a zadní nohy obou zvířat. Svalová tkáň byla uložena v samostatných sklenic a fixovaný ve formaldehydu, roztok 4%, pufrovaný (=roztoku formalinu 10%, pufrovaný)., Poté byla tkáň vložena do parafínu a nakrájena na 2 µm podle standardního provozního postupu (SOP RAH). Poté, snímky byly deparaffinized a běžně obarví hematoxylin a eosin (H&E) v automatické barvě stroje. Současně další snímky z 2 µm svalové tkáně, stejně jako pozitivní kontrolní vzorek svalové tkáně byly připraveny pro ruční barvení s „fosfowolframové kyseliny hematoxylin,“ zkráceně PTAH. Toto barvení je výhodné pro prokázání křížových pruhů kosterního svalu.,
šlechtitelské hodnoty a asociační analýza
v této studii jsme vyhodnotili 63 rysů používaných v šlechtitelském programu. Deregresované odhadované šlechtitelské hodnoty (DEBV) byly použity jako proměnná odezvy pro každou studovanou vlastnost. Odhadovaná hodnota chovu (EBV) všech hodnocených rysů byla deregresována pomocí metodiky popsané Garrickem et al. (2009). EBV každého zvířete bylo získáno z rutinního genetického hodnocení komerčním šlechtitelským programem (Topigs Norsvin) pomocí zvířecího modelu., Reliabilities za zvíře za účelem deregression byly získány z genetického hodnocení na základě metodiky Tier a Meyer (2004). Heritability použité pro deregresi byly také extrahovány z rutinního genetického hodnocení. Nakonec byly podle Garricka et al také odhadnuty váhové faktory založené na odhadované spolehlivosti DEBV. (2009) pomocí hodnoty 0,5 pro skalární c. K zajištění kvality DEBV, pouze zvířata s váhovými koeficienty větší než nula a spolehlivost DEBV větší než 0.,V asociačních analýzách bylo použito 20. Spolehlivost DEBV byla také získána podle Garrick et al. (2009).
asociační analýzy byly provedeny pomocí softwaru ASREML (Gilmour et al.,, 2009) použití následující lineární smíšené zvířecí model:
kde DEBVij je pozorován DEBV pro zvíře j, w je váhový faktor pro zbytkové, µ je celkový DEBV průměr populace, Ri je dopravcem stav (počet na škodlivé alely) 4 mutace já, aj je aditivní genetický efekt odhaduje pomocí pedigree založené na průměru matice vztahů, a zbytkových chyb. Asociace s hodnotou a-log10(hodnota P) větší než pět byly prohlášeny za významné.
výsledky
a 1.,5 Mb Segment na Chromozomu 6 Ovlivňuje Kojení Přežití u Prasat
Jsme analyzovali 31,638 zvířata z jednoho čistokrevného kance line (syntetické souladu s velké bílé pozadí), genotyp na Prasata 50K SNP čip (Sscrofa11.1 build) (Warre et al., 2019). Analýza ukázala, 1,5 Mb segment na chromozomu 6 (SSC6:48.75–50.25) ukazuje deficit v homozygotnosti spojené se sníženou laktaci, přežití (Tabulky 1 a 2). Haplotyp se ve sledované populaci segreguje se střední frekvencí alel 4,5% (9,0% nosné frekvence)., Haplotypová frekvence kolísá v posledním desetiletí, ale za poslední 3 roky se snížila (obrázek S1). Testovali jsme, zda byla frekvence poháněna heterozygotním výhodným efektem. Nicméně, našli jsme většinou negativní asociace s důležitými výběr rysů s výjimkou bederní hloubku a délku gestace (Tabulka 3), což naznačuje, že frekvence je čistě výsledkem genetického driftu.

Tabulka 1 SSC6 haplotyp vlastnosti.,

Tabulka 2 Dopravce od dopravce vrhy show 24% pokles laktace přežití ve srovnání s dopravcem-non-carrier vrhy. Významné výsledky jsou uvedeny tučně.

Tabulka 3 Vlastnosti významně spojené s heterozygotní přenašeči SPTBN4 odstranění.
vrhy 52 carrier-by-carrier (CxC) nevykazují významné snížení celkového počtu narozených nebo živých zvířat., Přežití laktace je však u vrhů CXC sníženo přibližně o 24% ve srovnání s maticemi carrier-by-noncarrier (CxN), což naznačuje, že homozygotní selata umírají během období laktace (Tabulka 2). Dále jsme zkoumali poznámky o čase a příčině úmrtnosti vrhů CxC. To odhalilo, že většina selat, která zemřela během prvních 24 hodin po narození. Většina těchto selat byla většinou označována zemědělci jako “ slabá selata při narození.,“
Celý-Sekvenování Genomu Analýza Odhaluje 16-bp Posunovými Smazání v SPTBN4 jako Pravděpodobné Kauzální Varianta
K identifikaci kauzální mutace, jsme zkoumal celý genom sekvence dat z 71 zvířata ze zkoumané populace a identifikovala pět dopravce zvířat. Vazebné nerovnováhy (LD) analýza ukázala, 267 SNP a indel variant ve vysoké LD (r2 > 0.8) s SSC6 haplotyp (Tabulka č. 1), většina je v perfektním LD (247 variant)., Pouze pět variant potenciálně ovlivňuje kódovací sekvenci (tři missense, jeden frameshift, jeden Splice-akceptor). Tři missense variant se předpokládá, že snášen SIFT (skóre > 0.18, Stolní S1), zatímco splice-akceptor varianta ovlivňuje gen kódující 28 bp peptidy neznámé funkce, je nepravděpodobné, že být kauzální. Nicméně, jedna varianta v naprosté LD (r2 = 1) s haplotyp byl předpovídán vysoký dopad; 16-bp posunovými delece v exonu 26 SPTBN4 gen (6:g.48801280delGACGGTGTACGCCGGT) (obrázky 1A, B)., Na posunovými smazání (ENSSSCP00000031537:p.Arg1902fs) přináší 30 nových aminokyselin a předčasnému stop kodonu, produkovat postižené a zkrácen spectrin beta non-erytrocytární protein 4 (SPTBN4). Mutantům chybí konečných 662 aminokyselin proteinu divokého typu (obrázek 1C), včetně domény pleckstrinové homologie (PH) potřebné pro transport bílkovin na membrány (Wang et al., 2018). Protein SPTBN4 je členem proteinů beta-spektrinu a je aktinem, který spojuje buněčnou membránu s aktinovým cytoskeletem., SPTBN4 mutace narušení cytoskeletální stroje ovládá správné lokalizace iontové kanály v myelinizovaných nervů, což způsobuje motorické neuropatie (Parkinson et al., 2001; Wang et al., 2018).

Obrázek 1 (a) genový model SPTBN4. Umístění postiženého 26. exonu je indikováno červeně. B) ilustrace výmazu 16 bp. Obrázek ukazuje divoký typ a mutantní exon. C) zarovnání sekvence proteinů mutantu (Mt) a divokého typu (Wt) sptbn4. Mutace indukuje 30 nových aminokyselin a předčasný stop kodon.,
Genotypizace Pět CxC Vrhy Potvrzuje SPTBN4 Smazání jako Pravděpodobný Viník
genotyp pět CxC vrhy pro 16-bp delece, který měl alespoň dvě selata (rozmezí 2-6), která zemřela během prvních 48 h po porodu. Pět vrhů vyrobilo 53 selat, z nichž 19 bylo homozygotních pro odstranění 16 bp (Tabulka 4). Všech 19 homozygotních selat zemřelo během 48 hodin po narození (18 do 24 hodin). Z 34 zbývajících selat (8 volně žijících druhů a 26 nosičů) zemřelo během 48 hodin pouze 1, pravděpodobně způsobené jinými (environmentálními) faktory.,

Tabulka 4 Genotypizace pravděpodobné kauzální 16-bp SPTBN4 posunovými smazání v pěti dopravce od dopravce vrhy. Součet na třídu genotypu je uveden tučně.
Selat Homozygotní pro SPTBN4 Smazání Trpí Myopatií a Zadní Končetiny, Ochrnutí
sledovali Jsme jedním z posledních CxC vrh (porodem datum: 28. dubna 2019), který produkoval šest zdravých, dva postižených (vzorky: 9912, 9916) (Obrázek 2A), a tři mrtvě narozených selat., Potvrdili jsme stav odstranění homozygotního sptbn4 u dvou postižených selat (tabulka S2). Kromě toho jsme u zdravých jedinců pozorovali čtyři heterozygotní nosiče a dvě homozygotní divoká selata. Jedno z mrtvě narozených selat (vzorek: 9921) bylo také homozygotní pro vymazání, zatímco ostatní dvě byly heterozygotní. Postižená selata trpí extrémní svalovou slabostí (obrázky 2B, C), paralýzou zadních končetin a třesem (Video S1). Proto selata nemohla chodit ani pít.,
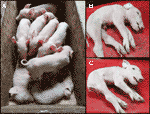
Obrázek 2 (A) Dva postižených selat (živý) spolu s šesti zdravých sourozenců. Selata pocházejí z jednoho CXC Páření farrowed dne 28. dubna 2019. B) postižené samčí selata 9912. C) postižená samička selata 9916.
Postižených Selat Nedostatek Křížové Rýhování v Hřbetní a Zadní Končetiny Kosterní Svaly,
Histopatologické vyšetření odhalilo rozptýlené degenerace svalových vláken v obou selat, a focally nekrózy a vaskulitidy u hřbetního svalu v jedné z selat (ID = 9912)., Navíc, fosfowolframové kyseliny hematoxylin (PTAH) barvení ukazuje odlišné zbarvení kosterních svalových vláken, což naznačuje snížení křížové rýhování, zejména v svaly hřbetní a zadní nohy postižených zvířat (Obrázek 3B), zatímco přední nohy se zdají být v pořádku (Obrázek 3A). Snížení křížových pruhů je indikováno abnormálním zbarvením a celkovou ztrátou objemu svalových vláken (obrázek 3B). Histopatologicky pozorované změny zadních nohou a hřbetních svalů svědčí o svalové dystrofii.,

Obrázek 3 (A) příčný řez kosterní sval z přední nohy. Černá šipka označuje normální zbarvení (tmavé) svalových vláken indikující přítomnost křížových pruhů. Ptah Bar = 50 µm. B) průřezový pohled na kosterní sval ze zadní nohy. Černá šipka označuje abnormální zbarvení (růžové) svalových vláken, což naznačuje nedostatek křížových pruhů. Žlutá šipka označuje normální zbarvení a přítomnost křížových pruhů. Ptah Bar = 50 µm.,
Diskuse
V této práci jsme zprávu román vrozená vada způsobující prasátko úmrtnosti pravděpodobné, vzhledem k 16 bp posunovými smazání v SPTBN4 gen. Selata trpí extrémní svalovou slabostí (myopatií) a zemřou během několika hodin po narození. Očekává se, že delece poskytne úplnou ztrátu funkce spektrin beta, non-erytrocytární 4 protein. SPTBN4 je členem rodiny spektrinu geny a je nutné pro iontový kanál clustering na ranvierovy zářezy, které ovlivňují akční potenciál (Devaux, 2010)., Mutace narušení cytoskeletální strojního zařízení, které ovládá správné lokalizace iontové kanály a funkce axonální domén hlavně na axon počáteční segmenty (AIS) a ranvierovy zářezy (Wang et al., 2018). Přesněji řečeno, postižená doména C-terminal SPTBN4 je rozhodující pro obchodování s kanálem kcnq2 a vzrušivost v uzlech Ranvier (Devaux, 2010).
následný následný výzkum identifikoval případy lidí a myší, které naznačovaly, že následný syndrom by pravděpodobně nebyl okamžitě smrtelný, ale spíše by způsobil těžkou myopatii., Podle údajů genotypu SNP se střední hustotou, které jsou k dispozici pro všechna zvířata v chovné populaci (N = 31,839), lze identifikovat nosiče. Mezi těmito nosiči byla prasnice, která byla v době identifikace přibližně v polovině těhotenství, utápěná kancem, který byl také nosičem. Chovná farma byla oznámena, aby dokumentovala vrh při narození. Pozorovaný fenotyp postižených selat (myopatie, zadní končetiny, ochrnutí, třes), byl zcela shodný s tím, co bylo pozorováno u lidských pacientů s homozygotní ztráta-of-funkce nebo složené heterozygotní mutace v SPTBN4 genu (OMIM: 606214)., Dva z lidských pacientů mají mutace ztrátové funkce v doméně PH (Wang et al., 2018), podporující, že ztráta domény PH u prasat by pravděpodobně vedla k úplné ztrátě funkce proteinu SPTBN4. U lidí podobné mutace vedou k těžké vrozené myopatii způsobené nepřítomností svalových vláken typu I, neuropatie a hluchoty (Knierim et al., 2017; Wang et al., 2018). Wang et al. (2018) také pozoroval motorickou axonální neuropatii u několika pacientů charakterizovaných vrozenou hypotenzí, hlubokou slabostí a ztrátou hlubokých reflexů šlach v raném dětství., Navíc nervové biopsie odhalily snížené uzlové kanály na + a žádné uzlové kanály kcnq2 k+, odhalující molekulární patologii způsobující dysfunkci nervového systému. Proto jsme dospěli k závěru, že tato varianta frameshift je pravděpodobnou kauzální mutací vedoucí k pozorovanému fenotypu a vyčerpání homozygotního genotypu v populaci. Budoucí studie by se mohly zaměřit na in vivo knockout genu SPTBN4 u prasat, podrobněji studovat syndrom a přidružený fenotyp.,
nepozorovali jsme degeneraci svalových vláken v předních nohách, zatímco svalová vlákna hřbetní a zadní nohy byla jasně ovlivněna. Toto pozorování by mohlo částečně vysvětlit paralýzu zadních končetin, zatímco přední nohy nejsou ovlivněny. Rozdíl mezi přední a zadní nohy svalová vlákna byla také popsána v chvějící myší, ve které SPTBN4 ztráta-off-funkce mutace způsobují motorická neuropatie, zadní končetiny, ochrnutí, třes, a centrální hluchota (Parkinson et al., 2001; Komada and Soriano, 2002). Parkinson et al., (2001) popisují snížené nervové vedení rychlosti v ischiatické nervy myší s chvějící alel způsobuje periferní zadní končetiny neuropatie. Exprese SPTBN4 u myší je omezena na mozek, míchu a sedací nervy a není pozorována v kosterním svalu, takže toto onemocnění je primárně neuronální defekt. Celkově zůstává nejasné, který mechanismus způsobuje nepřítomnost příznaků v předních končetinách. Tento „přírodní knockout“ u prasat může být užitečným zdrojem pro studium lidské nemoci, protože prasata jsou obvykle lepším modelem pro studium lidských onemocnění ve srovnání s druhy hlodavců., Kromě toho může být podrobněji studován důsledek ztráty funkce SPTBN4.
Efektivní velikost populace (Ne) studovaného plemene se odhaduje na přibližně 100 (Hidalgo et al., 2016). Při chovu zvířat nízké Ne zvyšuje riziko, že škodlivé alely náhodně stoupají. Předchozí studie navíc ukázaly, že recesivní letální alely mohou být poháněny výhodnými účinky u heterozygotů (Derks et al., 2018; Matika et al., 2019). Matika et al.,, 2019 našel recesivní stop-získané mutace v MSTN genu spojena s významným nárůstem svalové hloubka v heterozygoti. V naší studii však nenajdeme žádné důkazy o jakékoli heterozygotní výhodě. S aktuální genomické techniky můžeme nyní identifikovat škodlivé alely driftování na vyšší frekvence, a sledovat vznik nových škodlivé alely přesně, což umožňuje efektivnější čištění. Navíc výsledek tohoto typu studie výrazně zlepší vědomí „skrytých“ genetických defektů na úrovni chovatele i zemědělce., Bez jakýchkoli předchozích informací jsou vzácné vrozené vady často zaznamenány jako “ slabé selata.“A bez dalšího rozlišení specifických syndromů není další akce možná. Ve většině případů není známo, zda existuje genetický základ nebo že mohou existovat jiné matoucí účinky. S předchozími genomickými informacemi lze syndrom identifikovat ve srovnání s jinými případy a identifikovat nosiče, což vede k žalovatelným informacím.
úmrtnost na selata má velký hospodářský a sociální význam pro zvířata., Objev mutace SPTBN4 proto vedl k okamžité implementaci do šlechtitelského programu, aby se minimalizovala frekvence Páření nosiče po nosiči. To umožňuje vyhnout se narození postižených jedinců, čímž se zlepší dobré životní podmínky zvířat a sníží se ekonomické ztráty.
Závěr
V této studii jsme zprávu román vrozená vada pravděpodobně způsobena recesivní posunovými smazání v SPTBN4 genu u prasat. Nálezy jsou podporovány nápadnými podobnostmi se syndromovými fenotypy spojenými se SPTBN4 u lidí a myší., Studie umožňuje sledovat a očistit škodlivou alelu od populace. Nosič-nosič kříže může být zabráněno vyloučením postižených jedinců, čímž se sníží ekonomické ztráty a zlepšování životních podmínek zvířat. A konečně, tyto „přirozené knockouty“ získané v chovném průmyslu mohou poskytnout model pro lidské onemocnění a zvýšit naše chápání genové funkce v savčím clade a poskytnout potenciální model pro lidské onemocnění.,
Dostupnost Údajů Prohlášení
Etika Prohlášení
Etického přezkumu a schválení nebylo nutné pro zvíře studovat, protože data použitá v této studii byla získána jako součást rutinní sběr dat z Topigs Norsvin chovných programů, a ne speciálně pro účely tohoto projektu. Schválení Etické komise proto nebylo povinné. Sběr vzorků a záznam dat byly prováděny striktně podle nizozemského zákona o ochraně a dobrých životních podmínkách zvířat (Gezondheids – en welzijnswet voor dieren)., Písemný informovaný souhlas byl získán od vlastníků za účast jejich zvířat v této studii.
Autor Příspěvky
MG, H-JM, a MD koncipována a navržena studie. BH byl zodpovědný za obecnou organizaci a komunikaci s Topigs Norsvinem a zemědělci. MD A ML provedli analýzu dat. BD a KL prováděly laboratorní práce. SG-V provedla patologickou analýzu. MD napsal rukopis. H-JM, MG, BH, SG-V, BD, KL a ML poskytly užitečné připomínky a návrhy a pomohly navrhnout rukopis. Fenotypové údaje byly analyzovány ml., Všichni autoři přečetli a schválili konečný rukopis.
finanční Prostředky
Tento výzkum byl financován STW-Breed4Food Partnerství, projekt číslo 14283: Z posloupnosti fenotyp: detekce škodlivé odchylky od predikce funkčnosti. Tato studie byla finančně podpořena NWO-TTW a Breed4Food partnery Cobb Evropě, CRV, Hendrix Genetics a Topigs Norsvin. Kromě toho byla tato studie podpořena projektem IMAGE (Horizon 2020, č. 677353). Investoři neměli žádnou roli ve studijním designu, sběru a analýze dat, rozhodnutí o publikování nebo přípravě rukopisu., Použití clusteru HPC bylo umožněno společností CATAgroFood (sdílená výzkumná zařízení Wageningen UR).
Upozornění
data použitá v této studii byla získána jako součást rutinní sběr dat z Topigs Norsvin chovných programů, a ne speciálně pro účely tohoto projektu. Schválení Etické komise proto nebylo povinné. Sběr vzorků a záznam dat byly prováděny striktně podle nizozemského zákona o ochraně a dobrých životních podmínkách zvířat (Gezondheids – en welzijnswet voor dieren).,
Střet Zájmů
ML a BH jsou zaměstnanci Topigs Norsvin Výzkumné Centrum, výzkumný ústav úzce souvisí s jedním z investorů (Topigs Norsvin).
všichni zbývající autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
ostatní partneři Breed4Food Cobb Europe, CRV, Hendrix Genetics prohlašují, že pro tuto studii nemají konkurenční zájmy.,
autoři děkují franku van Haarenovi a Toonu Janssenovi za organizaci komunikace s farmou. Děkujeme Manon Houben za její příspěvek k patologickým vyšetřením. Děkujeme Mout Viller za pořízení videa a obrázků postižených a zdravých selat po narození. Děkujeme Jürgenu Harliziovi za péči o selata po předání. Děkujeme Gerdě van Eldikové za odběr vzorků. Děkujeme Egbert Knol a Egiel Hanenberg pro všeobecnou podporu a vedení od Topigs-Norsvin.,
doplňkový materiál
Video S1 / Video ukazující obě postižené osoby po narození.
tabulka S1 / genomická variace vysokého LD s haplotypem SSC6.
tabulka S2 / genotypizace vrhu CXC včetně dvou postižených jedinců (datum porodu: 28. dubna 2019). Tabulka ukazuje homozygotní stav del / del u dvou postižených selat 9912,9916.
tabulka S3 / mapování a statistiky pokrytí ze vzorků WGS.
obrázek S1 / SPTBN4 delece carrier frequency z let 2007-2019.