Bevezető
Ellentétben állatok mozgatni, hogy elkerüljék a kedvezőtlen környezetben, a kocsánytalan növények kiállítás egy magasan fejlett alkalmazkodás a bonyolult környezeti feltételek. Ennek a mély alkalmazkodóképességnek a eléréséhez a sejtek közötti kommunikációra van szükség. A sejt-sejt kommunikáció a növényekben robusztus intracelluláris jelátviteli feldolgozást és bonyolult intercelluláris jelátviteli hálózatokat foglal magában., Eddig legalább kilenc jelző anyagok nevű növényi hormonok, beleértve azonosítására, brassinosteroid (BR), cytokinin, gibberellins (GA), etilén, jasmonic sav (JA), strigolactone (SL), abszcizinsav esetében (ABA), valamint a szalicilsav (SA) fedezte fel (Druege et al., 2016; Verma et al., 2016). A genetikai és fiziológiai vizsgálatok feltárták e fenti hormonok kritikus szerepét és funkcionális mechanizmusait a növények növekedésében és fejlődésében (Gray, 2004)., A korábbi vizsgálatok alapján az auxin, BR, GA, SL és citokinin elsősorban a növény normál növekedése és fejlődése során működik, míg az ABA, etilén, JA és SA fontos szerepet játszik a növénynövekedés különböző biotikus és abiotikus stresszekre adott válaszában (Pieterse et al., 2009; Santner et al., 2009; Denance et al., 2013). Ezen hormonok némelyikének kettős szerepe van, például az ABA fontos szerepet játszik a magfejlesztésben és a nyugalomban is (Seo and Koshiba, 2002)., Bár egyes hormonok bizonyos szempontból meghatározó szerepet játszanak, sok hormon átfedésben van a tevékenységekkel, és a különböző hormonok kölcsönhatásai számos fejlődési szempontot és növekedést szabályoznak az endogén fejlődési és exogén jelekre adott válaszként.
az Auxin és a BR a növekedést elősegítő hormonok két fő osztálya., BR, egy csoport üzem-specifikus szteroid hormonok, amelyek kölcsönhatásba más phytohormones például azonosítására, cytokinin, etilén, GA, JA, meg SA, valamint szabályozza a sokféle növény növekedése, valamint a fejlődési folyamatokat, beleértve a csírázási, cella nyúlás, érrendszeri differenciálás, gázcserenyílások kialakulását, valamint a mozgás, virágzás, mind a férfi termékenységi (Saini et al., 2015). Érdekes, hogy ezen folyamatok mindegyikét az auxin is szabályozza, ami arra utal, hogy ez a két hormon kölcsönhatásban áll a növények fejlődésének szabályozásával., Ebben a felülvizsgálatban az auxin és a BR jelátvitelét vázoljuk fel a közelmúltban elért eredmények alapján, és áttekintjük az auxin és a BR közötti áthallást.
auxin jelátviteli útvonal
az Auxint először növényi hormonként ismerték fel, mivel szerepe volt a növényi tropizmusban a gravitációs vagy könnyű ingerekhez., Később azonosítására volt kémiailag azonosított indol-3-ecetsav, valamint kimutatták, hogy játszanak alapvető szerepet rengeteg növény fejlődési, illetve élettani folyamatok, beleértve embryogenesis, organogenesis, érrendszeri differenciálás, gyökér lőni fejlesztési, tropikus növekedés, valamint a gyümölcs-fejlesztési (Estelle, 2011).
az Arabidopsis genetikai elemzésével az auxin jelátvitel alapjául szolgáló molekuláris mechanizmust jól megvizsgálták. TRANSPORT INHIBITOR RESPONSE1 (TIR1) volt az auxin első azonosított nukleáris receptora (Ruegger et al., 1998; Dharmasiri et al., 2005)., A TIR1 az F-box fehérjéhez tartozó nukleáris fehérjét kódolja az SCF E3 ubiquitin ligáz fehérje komplex alegységeként (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005), a TIR1 mellett három további F-box fehérje van, nevezetesen az AUXIN jelző F BOX fehérjék (AFBs), amelyek auxin-kötő aktivitást mutatnak, és közvetítik az auxin jelátvitelt az Arabidopsis-ban (Badescu and Napier, 2006). A TIR1 receptor kölcsönhatásba léphet az AUX/IAA (auxin/indol-3-ecetsav) fehérjék (Dharmasiri et al., 2003)., Az AUX/IAA fehérjék az auxin jelátvitel negatív szabályozói, és az AUX / IAA 29 tagja van az Arabidopsis genomban kódolva. Az AUX / IAA fehérjék kölcsönhatásba léphetnek a transzkripciós szabályozók osztályával, az auxin válasz faktorokkal (ARF), hogy közvetítsék az auxinra adott transzkripciós válaszokat. Magas auxin szint mellett az AUX / IAA fehérjék kölcsönhatásba lépnek a TIR1-gyel az auxin coreceptoraként, és az SCFtir1 komplex mindenütt jelen lehet, így az ubiquitin–proteaszóma útvonalon (Gray et al., 2001; Lanza et al., 2012)., Amikor a pusztítás AUX/iaa-n repressors, az azonosítására transzkripciós szabályozók ARFs, amely tartalmazza 23 tagság lehet alól AUX/iaa-n elnyomás, így közvetíteni a azonosítására válaszok aktiválásával vagy az elnyomás a target gének (Guilfoyle közben, 2007). Az F-box protein és Aux/IAA vagy ARFs különböző készletei az auxin jelátalakítás (Goh et al., 2012; Guilfoyle, 2015; Salehin et al., 2015).,
az AUX/IAA transzkripciós represszorok és az ARF transzkripciós faktorok összehangolt működése összetett génszabályozó hálózatokat hoz létre, amelyekről a Physcomitrella (Lavy et al., 2016). A közelmúltban kiderült, hogy az SCF CULLIN1 (CUL1) alegysége kölcsönhatásba lép a TIR1-gyel, így szabályozza a TIR1 szubsztrátok stabilitását és az auxin jelátvitelt (Wang et al., 2016). A TIR1 és az AUX/IAA közötti kölcsönhatást az AUX/IAAs térbeli konformációja is befolyásolja, amelyet egy ciklofilin-izomeráz LRT2 szabályoz rizsben (Jing et al., 2015)., A 90-es HŐÜTÉSI tényező (HSP90) és az sgt1 társkísérője kölcsönhatásba lép a TIR1-gyel, és így szabályozza a TIR1 stabilitását, ami befolyásolja a TIR1 és az AUX/IAA és az auxin jelátvitel (Wang et al., 2016).
a TIR1-függő kanonikus auxin-jelátviteli út mellett a közelmúltban arról számoltak be, hogy az auxin nem kanonikus auxin-jelátviteli mechanizmuson keresztül sokféle fejlődési választ vált ki., Ebben a nem kanonikus auxin érzékelési folyamatban az ARF3 / ETTIN a génexpressziót folyamatspecifikus transzkripciós tényezőkkel való kölcsönhatások révén szabályozza, amelyek nagymértékben gazdagítják az auxin által közvetített növényi fejlődési sokféleséget (Simonini et al., 2016, 2017).
BR jelátviteli útvonal
a BRASSINOSZTEROIDOT először a pollenben fedezték fel a sejt megnyúlásának elősegítésére. Később kiderült, hogy a BR számos növénynövekedési szempontban játszik szerepet, és képes reagálni a biotikus és abiotikus stresszekre., Manapság a BR jelátviteli utat nagyrészt különböző módszerek kombinációi tisztázták, beleértve a molekuláris genetikát, a biokémiát, a proteomikát, a genomikát stb. A bri1 extracelluláris doménjéhez kötődő bri1 (BRI1) sejtfelszíni kináz ÉRZÉKETLENSÉG1 (BRI1) receptoraként azonosították, amely aktiválja kinázaktivitását, és így egy jel kaszkádra kapcsol a transzkripció szabályozására (Li és Chory, 1997; Wang et al., 2001; Kinoshita et al., 2005; Kim and Wang, 2010; Clouse, 2011; Hothorn et al., 2011; She et al., 2011; Oh et al., 2012)., A BRI1 a BRI1 észlelésekor kölcsönhatásba lép a bri1 társuló kináz 1 (BAK1) és homológ szomatikus EMBRIOGENEZISRECEPTOR Kinázokkal (SERKs), hogy aktívabb BR receptor komplexet (Li et al., 2002; Nam and Li, 2002; Wang et al., 2005; Tang et al., 2008; Gou et al., 2012). Aktivált BRI1 foszforilátok a plazmamembránhoz rögzített receptorszerű citoplazmatikus kinázok két szubsztrátja: BRASSINOSZTEROID-jelző KINÁZOK1 (BSK1) és konstitutív DIFFERENCIÁLNÖVEKEDÉS1 (CDG1) (Tang et al., 2008; Kim et al.,, 2011), amely viszont foszforilálja a BRI1-SZUPPRESSZOR1 (BSU1) nevű PP1-típusú foszfatázt a BSU1 aktiválásához, ami bsu1-foszforilációhoz és inaktiváláshoz vezet a GSK3-szerű kináz BRASSINOSZTEROID ÉRZÉKETLENSÉGE2 (BIN2). A BIN2 kináz aktivitását a hiszton-DEACETILÁZ HDA6 is gátolja, amely a BIN2 K189-ében kölcsönhatásba lép és deacetilál., Ha a BR-szintek alacsonyak, a BRI1 nyugalmi állapotban van a negatív szabályozója, a BRI1 kináz INHIBITOR 1 (BKI1) és a proteinfoszfatáz 2A (PP2A) miatt, míg a BIN2 foszforilát két br homológ transzkripciós faktor, a BRASSINAZOL RESISTANT1 (BZR1) és a BZR2 (más néven BES1 BRI1-EMS-szuppresszor 1) (He et al., 2002; Wang et al., 2002; Yin et al., 2002; Mora-Garcia et al., 2004; Kim et al., 2009, 2011; Kim és Wang, 2010). Ha a BR-szint magas, a BIN2 inaktiválódik, a BZR1 és a BZR2 pedig pp2a által defoszforilált, és a magba költözik, hogy megváltoztassa a BR-válasz gének ezreinek expresszióját (He et al.,, 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011).
A BR és az Auxin jelátvitel közötti szinergia
az Auxin és a BR jelútvonalak különböző szerepet játszanak, de a fejlődési folyamatok széles skáláján szinergikus és egymástól függő kölcsönhatásokat is mutattak. Például mind az auxin, mind a BR jelek elősegíthetik a sejtek terjeszkedését, és szinergikusan kölcsönhatásba léphetnek a hypocotyls megnyúlásának elősegítése érdekében (Nemhauser et al., 2004)., A hipokotil megnyúlás elősegítésében a két út egyikének válaszreakciója megköveteli a másik funkcióját, valamint a BR és auxin utak közötti kölcsönös függőséget (Nemhauser et al., 2004). Azonosítására fokozott hypocotyl hossza a vad típusú növények, de nem a BR-érzéketlen mutáns bri1-116, ez azonosítására-érzéketlen fenotípus bri1-116 elnyomták a domináns gain-of-function mutáns bzr1-1D, jelezve, BR vagy aktív BZR1 szükséges azonosítására támogatása hypocotyl nyúlás., Megállapítást nyert, hogy a BR jelátvitel a FITOKRÓM B4-3 (SOB3) SZUPPRESSZORÁVAL konvergál a sejtek megnyúlásának és hypocotyl növekedésének szabályozására az auxin által indukált kis AUXIN UP RNA19 (SAUR19) expresszió (Favero et al., 2017). Másrészről, az auxin szabályozott transzkripciós faktor kis szervméret 1 (SMOS1) a közelmúltban kiderült, hogy szabályozza a sejtek terjeszkedését a SMOS2/DLT-vel, a transzkripciós Co-szabályozók Gras családjának tagjával való közvetlen kölcsönhatás révén, amely pozitív szerepet játszik a BR jelátvitelben a rizsben (Kim et al., 2009; Tong et al., 2012; Hirano et al., 2017)., Azonosítására kapcsolódó mutánsok például iaa3, valamint arf6/arf8 kevésbé volt érzékeny BR volt, mint a vad-típusú hypocotyl nyúlás, s eltörölte a túlérzékenység bzr1-1D, hogy azonosítására, ami arra utal, hogy a BR valamint BZR1 támogatása hypocotyl nyúlás igényel ARF6/8. A genomra kiterjedő ChIP-Seq elemzés kimutatta, hogy az ARF6 számos genomikai célt (körülbelül 50%) oszt meg a BZR1-gyel és a fény/hőmérséklet által szabályozott PIF4 transzkripciós faktorral CHIP-Seq elemzésekkel (Oh et al., 2014)., A bzr1 és a PIF4 kölcsönhatásba lép az ARF6-tal és aktiválja a közös célgéneket azáltal, hogy a hypocotyls megnyúlás során együttműködően kötődik a megosztott célgénekhez (Oh et al., 2014) és ezen átfedő célgének nagy része a sejttágulásban részt vevő sejtfalfehérjéket kódolja.
Brassinosteroid és auxin szintén fontos szerepet játszanak a gyökér apikális merisztem (RAM) (Durbak et al., 2012). A RAM áll egy kis csoport ritkán osztódó sejtek úgynevezett nyugalmi központ (QC), körülvéve őssejtek, hogy ad okot, hogy a különböző toot szövet típusok., A gyökér őssejtpopuláció fenntartását a WUSCHEL-rel kapcsolatos HOMEOBOX 5 (Wox5) (Sarkar et al., 2007). A wox5 az auxin jelzéssel a QC-re korlátozódik, és elősegíti a PLT gének megfelelő expresszióját (Aida et al., 2004; Ding és Frml, 2010). A BR-receptor gén BRASSINOSZTEROID érzéketlen 1 (BRI1) mutációi aberráns sejtciklus-progressziót eredményeznek a RAM-ban, és kisebb RAM-ot okoznak (Gonzalez-Garcia et al., 2011; Hacham et al., 2011). Az auxinról ismert, hogy stimulálja a BR bioszintézisét (Chung et al.,, 2011), de a BR aktivitása nem befolyásolja a PIN gének expresszióját (Hacham et al., 2011). A BR mutánsok gyökércsúcs fenotípusai nem mutatnak ugyanazt, mint az auxin mutánsok (Gonzalez-Garcia et al., 2011), jelezve, hogy BR törvény a RAM függetlenül auxin.
Brassinosteroid és auxin jelek szinergikusan szükségesek a vaszkuláris kötegek radiális mintaképzésében (Ibanes et al., 2009)., A matematikai modellezés és biológiai kísérletek kombinációjával az aszimmetrikus auxin poláris transzport által létrehozott auxin maxima, de az auxin szinteken nem bekövetkező változások fontosak a vaszkuláris kötegek pozicionálásához. Kimutatták, hogy a BR jel elősegítő jelként szolgál a provaszkuláris gyűrűben lévő sejtek számára, amelyek összhangban vannak az auxin maximával. Így az érrendszeri kötegek időszakos elrendezésének megteremtése a hajtásban e két növényi hormon (Ibanes et al., 2009)., Mindkét jel a növénygyökér fejlődésében is szerepet játszik, és a BR és auxin kölcsönhatását a brevis RADIX (BRX) közvetíti e folyamat során. BRX fontos a sebességkorlátozó bioszintézis BR és BR exogén alkalmazás képes megmenteni brx mutáns hibák. Továbbá azonosítására reagál a génexpresszió világszerte csökkent a brx mutáns, a kifejezés pedig a BRX erősen által kiváltott azonosítására, illetve elnyomott a BR, ami BR bioszintézis, illetve azonosítására jelzés keresztül kapcsolódnak a visszacsatolás érintő BRX a gyökér fejlődése (Mouchel et al., 2006).,
A Brassinoszteroidok és az auxin szintén szinergikus szerepet játszanak az oldalirányú gyökérfejlődés során. A BRs elsősorban az oldalsó gyökér primordia iniciációjában működik, míg az auxin szükséges mind az oldalirányú gyökérképződés megindításához, mind megjelenési szakaszaihoz (Casimiro et al., 2001; Bhalerao et al., 2002; Benkova et al., 2003; Bao et al., 2004). E folyamatok során a BRs növeli az LRP indítását azáltal, hogy elősegíti az acropetal auxin transzportot a gyökérben, de nem befolyásolja az endogén IAA szintet (Bao et al., 2004)., Mindezek a jelentések arra utalnak, hogy a BR és az auxin közötti keresztút fontos szerepet játszik a növénynövekedés és-fejlődés szabályozásában.
BR Szabályozza Azonosítására Jelzés
Mellesleg a kölcsönös együttműködés a azonosítására, valamint BR jelek során a növény fejlődését, BR kitalálhatunk azonosítására jel útja, több szinten. BZR1 együttműködik ARF fehérjék közvetlenül a cél több azonosítására jelzés összetevők kialakulásában szerepet játszó gének azonosítására anyagcsere-például közlekedési jelzés, beleértve AUX/iaa-n, Csapok, TIR1, valamint ARFs, stb. (Sun et al., 2010)., Megállapítást nyert, hogy az AUX / IAA fehérjék részt vesznek a BR-válaszokban, az iaa7 / axr2-1 és az iaa17 / axr3 – 3 mutánsok pedig aberráns BR-érzékenységet és aberráns BR-indukált gén expressziót mutattak szervfüggő módon (Nakamura et al., 2006). Az exogén brassinolid (BL) kezelés indukálhatja az auxinra reagáló gének expresszióját, például IAA5, IAA19, IAA17 stb. a fenti gének expresszióját a BR bioszintetikus mutáns de-etiolated2 (det2) szabályozza, ami azt jelzi, hogy a funkcionális BR bioszintézis részben szükséges az auxin-függő gén expressziójához (Nakamura et al.,, 2003; Kim et al., 2006). Ezenkívül a BR az auxin-áramlást is befolyásolja az auxin-exportőrök expressziójának szabályozásával, mint például a PIN4 és a PIN7 (Nakamura et al., 2004). A növényi gravitációs válaszok során a BRs fokozhatja az auxin exportőr PIN2 poláris felhalmozódását a gyökér merisztémzónában, és ezáltal befolyásolhatja az auxin redisztribúcióját a gyökércsúcstól a nyúlási zónák felé, és az IAA szintek különbségét eredményezi a gyökerek felső és alsó oldalán a növényi gravitropizmus kiváltására., E folyamat során a BR aktivált ROP2 fontos szerepet játszik a PIN2 funkcionális lokalizációjának modulálásában az F-aktinok összeszerelésének/összeszerelésének szabályozásával (Li et al., 2005). További vizsgálatok azt mutatták, hogy a csökkent BL-észlelés és / vagy-koncentráció indukálhatja a cyp79b2-t, a triptofánt indol-3-acetaldoximmá konvertáló enzimet kódoló gént, amely így befolyásolja az eloszlást (Kim et al., 2007).
ezenkívül azt találták, hogy a BR jel szabályozhatja az auxin jelátvitelt a negatív szabályozó gsk3 kinase BIN2 által., Az arf2 auxin válaszfaktort BIN2 kölcsönhatásban lévő fehérjeként azonosították egy élesztő két-hibrid szűrőben, és a kináz-teszt azt mutatta, hogy a BIN2 képes foszforilálni az ARF2-t. Az ARF2 foszforilációja a DNS-kötő képességének elvesztését és a célgének elnyomó aktivitását eredményezi (Vert et al., 2008). Az ARF2 egy bzr1 célgén, kifejeződését a BR-kezelés csökkenti (Sun et al., 2010)., Ezenkívül a BIN2 foszforilálhatja az ARF7-et és az ARF19-et, hogy elnyomja az AUX/IAAs-szal való kölcsönhatásukat, és ezáltal fokozza a transzkripciós aktivitást célgénjeiken oldalsó SZERVHATÁROK-DOMAIN16 (LBD16) és LBD29 az oldalsó gyökér organogenezis szabályozására (Cho et al., 2014). A BR azonban kisebb szerepet játszik ebben a folyamatban, és a BIN2 a TRACHEÁRIS elem differenciálódási gátló faktor (TDIF)–TDIF RECEPTOR (TDR) modul (Cho et al., 2014). Együtt, BR szabályozhatja auxin reponses befolyásolása révén különböző auxin jelző alkatrészek.,
az Auxin szabályozza a BR jelátvitelt
másrészt az auxin bizonyos szempontból is szabályozhatja a BR jelútját. A DWARF4 expressziója, amely a BR bioszintézis szempontjából döntő fontosságú hidroxiláz az endogén BR szint szabályozásához, az auxinfüggő. Az Auxin-kezelés észrevehetően stimulálhatja a DWARF4 expresszióját, az auxin pedig gátolhatja a BZR1 kötődését a DWARF4 promoteréhez. A DWARF4 auxin általi indukciója auxin jelátviteli utat igényel, de nem BR jelátviteli utat (Chung et al., 2011; Josimitsu et al., 2011)., CPD katalizáló C-3 oxidációja BR aktiválta BRX, egy feltételezett transzkripciós faktor ható downstream auxin jelző (Mouchel et al., 2006). A rizs további vizsgálata azt jelzi, hogy az exogén auxin fokozhatja a BR receptor gén OsBRI1 transzkripciós expressziós szintjét, ami arra utal, hogy az auxin a BR receptorok szabályozásával fokozza a BR jelátvitelt (Sakamoto et al., 2013). Ezenkívül az OsBRI1 promoter rendelkezik egy upstream auxin-response element (auxre) motívummal, amelyet ARF transzkripciós tényezők céloznak meg., Ezenkívül a mutáns vizsgálatok azt mutatják, hogy az Auxre mutációja esetén az OsBRI1 expressziójának auxin általi indukciója megszűnik, valamint az OsBRI1 expresszióját az arf mutánsban (Sakamoto et al., 2013). Arról számoltak be, hogy az OsARF19 kötődik az OsBRI1 promoteréhez, és pozitívan szabályozza annak kifejeződését, amely aktiválja a BR jelátvitelt (Zhang et al., 2015). BES1 kötődnek a szervező a KIS AZONOSÍTÁSÁRA-FEL-RNS 15 (SAUR15) közvetítőnek BR korai válasz gén Osztályába, pedig ez kötelező is lehet fokozni, az azonosítására kezelés (Walcher, valamint Nemhauser, 2012)., Együttesen az auxin hatással lehet A BR-re és a BR-re is.
Záró megjegyzések és jövőbeli perspektíva
az elmúlt közel négy évtizedben az auxin-BR útvonal kölcsönhatásokkal kapcsolatos tanulmányok egyre több kutató érdeklődését vonzották. A fiziológiai, molekuláris, genetikai és biokémiai eszközök alkalmazása nagymértékben elmélyítette a kérdés megértését., A korábbi vizsgálatok alapján a BR és az auxin szinergikusan vesz részt több Növényfejlődési folyamatban, többek között: hipokotil nyúlás, vaszkuláris kötegfejlődés, gyökérfejlődés és tropizmusok stb. A kölcsönös együttműködés a azonosítására, valamint BR bonyolult benne, hogy számos folyamatok molekuláris szintű megosztása révén, ugyanazon cél gének, szabályozó egymást kölcsönösen több szinten (1.Ábra).
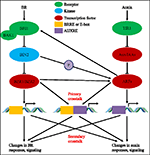
1.ábra. Az auxin-brassinosteroid (BR) crosstalk modellje., Az Arabidopsisban a BR és az auxin jel érzékelését a BRI1, illetve a TIR1 receptorok ismerik fel. A BR kötődik a BRI1 extracelluláris doménjéhez, és elősegíti, hogy kölcsönhatásba lépjen a bak1 co-receptorral, hogy aktívabb BR-receptor komplexet képezzen, ami viszont a BIN2 dezfoszforilációjához és inaktiválásához vezet. A BIN2 inaktiválása két BR homológ transzkripciós faktor, a bzr1 és a bzr2 defoszforilációjához vezet, amelyek a nucleusba költöznek, hogy aktiválják a brre-t vagy az E-boxot tartalmazó gének transzkripcióját promoter régiójukban., A BIN2 képes foszforilálni az ARF7-et és az ARF19-et is, hogy elnyomja az AUX/IAAs-szal való kölcsönhatásukat, és ezáltal fokozza a transzkripciós aktivitást célgénjeiken. TIR1 az auxin jelátvitelt fogadja, és az auxin társreceptoraként kölcsönhatásba lép az AUX/IAA fehérjékkel. Az AUX/IAA ezután az ubiquitin–proteaszóma úton lebomlik, és az auxin transzkripciós szabályozók auxin válasz faktorok (ARFs) felszabadulnak az AUX / IAA elnyomásból, és aktiválják a gének transzkripcióját auxin reagáló elemekkel (auxre) szabályozó régiójukban., Egyes ARF-ek a BRI1 promóteréhez is kötődnek, és pozitívan szabályozzák annak kifejeződését, amely ezután aktiválja a BR jelátvitelt. Az elsődleges áthallás olyan gének aktiválásával történik, amelyek mind a brre/E-boxot, mind az auxre-t tartalmazzák promoter régiójukban, lehetővé téve mindkét jelátviteli út számára a transzkripció közvetlen szabályozását. Másodlagos áthallás fordul elő a kifejezés, a gének, amelyek vagy azonosítására, vagy BR érzékeny, de a tevékenységek, amelyek ellenőrzése kifejezést a gének szabályozzák a válasz jelzés más hormonok.,
Foszforiláció rendelet döntő szerepet játszik BR jelátvitelben, főleg az észlelés folyamata, BR érzékelhető keresztül BRI1 kináz receptor, valamint BAK1 kináz co-receptorok, végül ellenőrzések BR szabályozott gén kifejeződése keresztül befolyásoló downstream transzkripciós faktorok, mint például BES1/BZR1 tevékenységek (Ő et al., 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011). Az ubiquitinációs szabályozás azonban elengedhetetlennek tűnik az auxin jelzéshez., Egyszer azonosítására kötődik TIR1 receptor, amely működik, mint egy ubiquitin E3-ligase, az aktivált TIR1 E3-ligase ubiquitinates AUX/iaa-n fehérjék, vezet a bomlás ezek repressors, de-elnyomja az ARF transzkripciós faktorok, végül okok azonosítására szabályozott gén expressziós mintázat megváltozik, illetve növekedési válaszok (Gray et al., 1999, 2002; Hellmann et al., 2003; Quint et al., 2005). Mivel megállapítást nyert, hogy a BR jelátvitelben jól ismert BIN2 kináz foszforilálhatja és fokozhatja az ARF-ek, például az ARF2 és az ARF7 (Vert et al., 2008; Cho et al.,, 2014), érdekes lesz megvizsgálni, hogy az olyan kinázok, mint a BIN2, amelyek részt vesznek a BR jelzésben, kölcsönhatásba léphetnek-e más auxin jelző komponensekkel is, mint például a TIR1 receptor vagy az AUX/IAA kompresszorok, és befolyásolhatják a TIR1 E3-ligáz aktivitást vagy az AUX/IAA fehérje stabilitásokat. Másrészt foglalkozni kell az ubiquitináció szerepével a BR jelátvitelben is, különösen akkor, ha a TIR1 E3-ligáz közvetlenül kölcsönhatásba léphet a BR jelátviteli komponensekkel, és szabályozhatja fehérje-stabilitásukat.,
emellett az auxin response DR5 és más auxin riporterek alkalmazásával megfigyelték, hogy az auxin szöveti vagy sejtfüggő módon szabályozza a növények növekedését és fejlődését. A különböző transzkripciós kimenetek a celluláris és környezeti kontextustól függően (Clark et al., 2014; Etchells et al., 2016; Lavy et al., 2016)., Bár az eső BR jelzés kimutatták, hogy ellenőrizzék a root növekedés az antagonisztikus a azonosítására (Chaiwanon, valamint Wang, 2015-ben), még ismeretlen, ha a szövet vagy sejt BR jelzés, ami láthatóvá pBZR1:BZR1-YFP, az is fontos, hogy ellenőrizzék, hogy más folyamatok mellett a gyökér fejlődése. Ezenkívül jelenleg fluoreszcens aktivált sejtválogatással vagy lézeres mikrodiszekcióval, nagyfelbontású génexpressziós elemzéssel kombinálva lehetséges az auxin és a BR eloszlások részletes szövet-vagy pincetérképének elkészítése., Ez végül vezet a cím, ha az auxin crosstalks a BR egy szövet vagy sejtes módon.
szerzői hozzájárulások
minden szerző részt vett a felülvizsgálati kézirat írásában. További információ a szerzőségről, kérjük, olvassa el a szerzői irányelveinket.,
finanszírozás
ezt a munkát a kínai Nemzeti Természettudományi Alapítvány (31500229 és 31770305 projektek), Shandong tartomány Természettudományi alapítványa (2017c03), Shandong tartományi alapok kiváló fiatal tudósok számára (2014jq201408) támogatja.
összeférhetetlenségi nyilatkozat
a szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségnek tekinthetők.
Clouse, S. D. (2011)., Brassinosteroid jelátvitel: a receptor kináz aktiválásától a transzkripciós hálózatokig, amelyek szabályozzák a növény fejlődését. Növényi Sejt 23, 1219-1230. doi: 10.1105 / tpc.111.084475
PubMed Abstract | CrossRef teljes szöveg | Google Scholar
Estelle, M. (2011). Auxin jelzés: a Szintézistől a Rendszerbiológiáig; Tárgygyűjtemény a Cold Spring Harbor Perspektíváiból a biológiában. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Google Scholar
Guilfoyle, T. J. (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar