introduktion
Till skillnad från djur som kan röra sig för att undvika de negativa omgivningarna uppvisar sessila växter en högt utvecklad anpassning till de komplicerade miljöförhållandena. För att uppnå denna djupa anpassningsförmåga är kommunikation mellan celler nödvändig. Cell till cellkommunikation i växter innebär robust intracellulär signalbehandling och invecklade intercellulära signalnätverk., Hittills finns det minst nio signalämnen, namngivna växthormoner, inklusive auxin, brassinosteroid (BR), cytokinin, gibberelliner (GA), etylen, jasmonsyra (JA), strigolakton (SL), abscisinsyra (ABA) och salicylsyra (SA) upptäckt (Druege et al., 2016; Verma et al., 2016). De genetiska och fysiologiska studierna har avslöjat de kritiska rollerna och funktionella mekanismerna för dessa ovanstående hormoner i växttillväxt och utveckling (Gray, 2004)., Baserat på tidigare studier fungerar auxin, BR, GA, SL och cytokinin huvudsakligen under normal växttillväxt och utveckling, medan ABA, etylen, JA och SA spelar viktiga roller i växttillväxtsvar på olika biotiska och abiotiska påfrestningar (Pieterse et al., 2009; Santner et al., 2009; Denance m.fl., 2013). Och även några av dessa hormoner har dubbla roller, till exempel spelar ABA också viktiga roller i fröutveckling och dormancy (Seo och Koshiba, 2002)., Även om varje hormon spelar dominerande roller i vissa aspekter, har många hormoner överlappade aktiviteter och interaktionerna mellan olika hormoner kontrollerar många utvecklingsaspekter och tillväxt som svar på endogena utvecklings-och exogena signaler.
Auxin och BR är två huvudklasser av tillväxtbefrämjande hormoner., BR, en grupp av växtspecifika steroidhormoner som kan interagera med andra fytohormoner som auxin, cytokinin, eten, GA, JA och SA och reglera ett brett spektrum av växttillväxt och utvecklingsprocesser inklusive frögroning, cellförlängning, vaskulär differentiering, stomatabildning och rörelse, blommande och manlig fertilitet (Saini et al., 2015). Intressant, var och en av dessa processer styrs också av auxin, vilket tyder på dessa två hormoner samspel för att kontrollera växtutveckling., I den här översynen kommer vi att skissera signaltransduktionen av auxin och BR baserat på de senaste framstegen och se över korspalken mellan auxin och BR-medierad växttillväxt och utveckling.
Auxin Signaleringsväg
Auxin erkändes först som växthormon på grund av dess roll i växt tropism till gravitation eller ljusstimuli., Senare auxin identifierades kemiskt som indol-3-ättiksyra och visade sig spela viktiga roller i överflöd av växtutvecklings-och fysiologiska processer, inklusive embryogenes, organogenes, vaskulär differentiering, rot-och skottutveckling, tropisk tillväxt och fruktutveckling (Estelle, 2011).
med hjälp av genetisk analys i Arabidopsis har den molekylära mekanism som ligger till grund för auxin-signaltransduktionen undersökts väl. TRANSPORT-HÄMMARE RESPONSE1 (TIR1) var den första identifierade nukleär receptor av auxin (Ruegger et al., 1998; Dharmasiri m.fl., 2005)., TIR1 kodar för ett kärnprotein som tillhör f-boxproteinet som en subenhet av SCF E3 ubiquitinligasproteinkomplex (Gray et al., 1999, 2002; Hellmann m.fl., 2003; Quint et al., 2005), I tillägg till TIR1, finns det ytterligare tre F-box proteiner nämligen AUXIN SIGNALERING F BOX Proteiner (AFBs) som visar auxin-bindande verksamhet och förmedla auxin signalering i Arabidopsis (Badescu och Napier, 2006). TIR1 receptor kan interagera med en grupp av AUX/IAA (auxin/indol-3-ättiksyra) proteiner (Dharmasiri et al., 2003)., AUX / IAA proteiner är negativa regulatorer av auxin signalering och det finns 29 medlemmar av AUX/IAA kodad i Arabidopsis genom. AUX / IAA-proteiner kan interagera med klassen av transkriptionella regulatorer, auxin response factors (ARF), för att medla transkriptionella svar på auxin. Under hög auxin nivå, AUX/IAA proteiner interagerar med TIR1 som coreceptor av auxin, och kan vara ubiquitinated av SCFtir1 komplexa och därmed försämras genom ubiquitin–proteasom väg (Gray et al., 2001; Lanza et al., 2012)., Vid förstörelsen av AUX/IAA repressorer, auxin transcriptional regulatorer ARFs som innehåller 23 medlemskap kan släppas från AUX / IAA repression och därmed medla auxin svar genom aktivering eller förtryck av målgener (Guilfoyle och Hagen, 2007). Olika uppsättningar av F-box protein och AUX/IAA eller ARFs sluta komplexitet under auxin signaltransduktion (Goh et al., 2012; Guilfoyle, 2015; Salehin et al., 2015).,
den samordnade åtgärden av Aux / IAA-transkriptionella repressorer och arf-transkriptionsfaktorer producerar komplexa genreglerande nätverk som också rapporterades i Physcomitrella (Lavy et al., 2016). Nyligen, det upptäcktes att CULLIN1 (CUL1) subenheten av SCF interagerar med TIR1 och reglerar därför TIR1 substrat stabilitet och auxin signalering (Wang et al., 2016). Samspelet mellan TIR1 och Aux/IAA är också påverkad av den rumsliga gestaltning av Aux/IAAs, som kontrolleras av en cyclophilin isomerase LRT2 i ris (Jing et al., 2015)., HEAT SHOCK FAKTOR 90 (HSP90) och co-förkläde SGT1 respektive interagerar med TIR1 och reglerar därför TIR1 stabilitet, vilket påverkar samspelet mellan TIR1 och Aux/IAA och auxin signalering (Wang et al., 2016).
förutom den tir1-beroende kanoniska auxin-signaleringsvägen har auxin nyligen rapporterats framkalla ett varierat utbud av utvecklingssvar genom en icke-kanonisk auxin-signaleringsmekanism., I denna icke-kanoniska auxin-avkänningsprocess kontrollerar ARF3/ETTIN genuttryck genom interaktioner med processspecifika transkriptionsfaktorer, vilket i hög grad berikar auxinmedierad växtutvecklingsdiversitet (Simonini et al., 2016, 2017).
BR Signaleringsväg
BRASSINOSTEROID upptäcktes först i pollen för dess förmåga att främja cellförlängning. Senare konstaterades att BR spelar roller i ett brett spektrum av växttillväxtaspekter och kan reagera på biotiska och abiotiska påfrestningar., Numera BR signal transduction pathway klargjordes till stor del genom kombinationer av olika metoder, inklusive molekylär genetik, biokemi, proteomik, och genomik, etc. Cell-ytan kinas BRASSINOSTEROID INSENSITIVE1 (BRI1) identifierades som receptorn BR som kan binda till den extracellulära domänen av BRI1 och aktivera dess kinas aktivitet och därmed slå på en signal kaskad att reglera transkription (Li och Chory, 1997; Wang et al., 2001; Kinoshita m.fl., 2005; Kim och Wang, 2010; Clouse, 2011; Hothorn et al., 2011; Hon et al., 2011; O, et al., 2012)., Vid perception av BR, BRI1 interagerar med co-receptor BRI1-ASSOCIERADE KINASE 1 (BAK1) och dess homolog SOMATISK EMBRYOGENESISRECEPTOR Kinaser (SERKs) för att bilda en mer aktiv BR receptor-komplex (Li et al. 2002; Nam och Li, 2002; Wang m.fl., 2005; Tang et al., 2008; Gou m.fl., 2012). Aktiveras BRI1 phosphorylates två substrat av plasma membranet förankrade receptor-som cytoplasmisk kinaser: BRASSINOSTEROID-SIGNALERING KINASES1 (BSK1) och GRUNDLÄGGANDE DIFFERENTIAL TILLVÄXT1 (CDG1) (Tang et al., 2008; Kim et al.,, 2011), vilket i sin tur phosphorylates en PP1-typ fosfatas som heter BRI1-SUPPRESSOR1 (BSU1) för att aktivera BSU1, vilket leder till BSU1 dephosphorylation och inaktivering av GSK3-som kinas BRASSINOSTEROID INSENSITIVE2 (BIN2). Bin2-kinasaktiviteten hämmas också av HISTONDEACETYLAS HDA6, som interagerar och deacetylater vid BIN2: s K189., När BR-nivåer är låga, BRI1 är vilande på grund av dess negativa regulator, BRI1 KINASE INHIBITOR 1 (BKI1) och protein fosfatas 2A (PP2A), medan BIN2 phosphorylate två BR homolog transkriptionsfaktorer, BRASSINAZOLE RESISTANT1 (BZR1) och BZR2 (som också heter BES1 för BRI1-EMS-SUPPRESSOR 1) (Han et al., 2002; Wang et al., 2002; Yin et al., 2002; Mora-Garcia et al., 2004; Kim et al., 2009, 2011; Kim och Wang, 2010). När BR-nivåerna är höga inaktiveras BIN2, och BZR1 och BZR2 defosforyleras av PP2A och flyttas in i kärnan för att förändra uttrycket av tusentals BR-gener (he et al.,, 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011).
synergin mellan BR Och Auxin-signalering
Auxin-och BR-signalvägar spelar olika roller, men de visade också synergistiska och ömsesidiga interaktioner i ett brett spektrum av utvecklingsprocesser. Till exempel kan både auxin-och BR-signaler främja cellutvidgning och interagera synergistiskt för att främja hypokotyls förlängning (Nemhauser et al., 2004)., Svaret på en av de två vägarna för att främja hypokotylförlängning kräver funktionen hos den andra och det ömsesidiga beroendet mellan BR och auxin vägar (Nemhauser et al., 2004). Auxin ökade hypokotyllängden i vilda växter men inte i BR-okänslig mutant bri1-116, och denna auxin-okänslig fenotyp av bri1-116 undertrycktes av den dominerande förstärkningen-av-funktion mutant bzr1-1D, vilket indikerar BR eller aktiv BZR1 krävs för auxin främjande av hypokotyl förlängning., Det har konstaterats att BR signalering konvergerar med SUPPRESSOR AV PHYTOCHROME B4-3 (SOB3) för att kontrollera cell töjning och hypocotyl tillväxt genom förordning av auxin inducerad SMÅ AUXIN UPP RNA19 (SAUR19) uttryck (Favero et al., 2017). Å andra sidan, auxin reglerade transkriptionsfaktor LITEN ORGEL STORLEK 1 (SMOS1) har nyligen visat att styra cellens expansion genom direkt interaktion med SMOS2/DLT, en medlem av GRAS familj på transkriptionell co-regulatorer som spelar en positiv roll i BR signalering i ris (Kim et al., 2009; Lee et al., 2012; Hirano et al., 2017)., Auxin relaterade mutanter som iaa3 och arf6/arf8 var mindre känsliga för BR än var vild-typ för hypocotyl töjning och avskaffat överkänslighet av bzr1-1D till auxin, vilket tyder på BR och BZR1 främjande av hypocotyl töjning kräver ARF6/8. Genomet-wide ChIP-Seq-analysen visade att ARF6 delar ett stort antal genomiska mål (cirka 50%) med BZR1 och ljus/temperaturreglerade transkriptionsfaktor PIF4 av CHIP-Seq-analyser (Oh et al., 2014)., Bzr1 och PIF4 interagerar med ARF6 och aktiverar Delade målgener genom att binda till delade målgener kooperativt under hypokotylsförlängning (Oh et al., 2014) och många av dessa överlappande målgener kodar cellväggsproteiner som är involverade i cellutvidgning.
Brassinosteroid och auxin spelar också viktiga roller i underhållet av root apical meristem (RAM) (Durbak et al., 2012). RAM består av en liten grupp av sällan delande celler som kallas quiescent center (QC), omgiven av stamceller som ger upphov till de olika toot vävnadstyperna., Underhåll av rot stamceller befolkningen är reglerad genom WUSCHEL-RELATERADE HOMEOBOX 5 (WOX5) (Sarkar et al., 2007). WOX5 är begränsad till QC genom auxin signalering och underlättar korrekt uttryck för PLT gener (Aida et al., 2004; Ding och Friml, 2010). Mutationer i BR-receptorgenen BRASSINOSTEROID okänslig 1 (BRI1) resulterar i avvikande cellcykelprogression i RAM och orsaka en mindre baggar (Gonzalez-Garcia et al., 2011; Hacham m.fl., 2011). Auxin är känt för att stimulera biosyntesen av BR (Chung et al.,, 2011), men aktiviteten hos BR påverkar inte uttrycket av PIN-gener (Hacham et al., 2011). Rotspetsfenotyper av BR-mutanter visar inte detsamma som auxin-mutanterna (Gonzalez-Garcia et al., 2011), vilket indikerar att BR agerar på RAM oberoende av auxin.
Brassinosteroid-och auxinsignaler krävs också synergistiskt i den radiella mönsterbildningen av vaskulära buntar (Ibanes et al., 2009)., Genom kombinationerna av matematisk modellering och biologiska experiment är auxin maxima, etablerad av asymmetrisk auxin polär transport, men inte förändringar på auxinnivåer viktiga för positionering av kärlbuntarna. BR-signalen visades fungera som en främjande signal för antalet celler i den provaskulära ringen som överensstämmer med auxin maxima. Således inrättandet av periodiska arrangemang av vaskulära buntar i skottet är under den samordnade verkan av dessa två växthormoner (Ibanes et al., 2009)., Båda signalerna är också involverade i växternas rotutveckling och samspelet mellan BR och auxin förmedlas av BREVIS RADIX (BRX) under denna process. BRX är viktigt för den hastighetsbegränsande biosyntesen av BR och BR exogen applikation kan rädda brx mutantfel. Dessutom är auxin-responsivt genuttryck globalt nedsatt i brx-mutant, och uttrycket av BRX induceras starkt av auxin och undertrycks av BR, vilket innebär att BR-biosyntes och auxin-signalering är anslutna genom en återkopplingsslinga som involverar BRX under rotutveckling (Mouchel et al., 2006).,
Brassinosteroider och auxin spelar också synergistiska roller under lateral rotutveckling. BRs fungerar huvudsakligen vid lateral rot primordia initiering medan auxin krävs för både inledande och uppkomststadier av lateral Rotbildning (Casimiro et al., 2001; Bhalerao m.fl., 2002; Benkova et al., 2003; Bao m.fl., 2004). Under dessa processer, BRs öka LRP initiering genom att främja acropetal auxin transport i roten men inte genom att påverka endogena IAA nivå (Bao et al., 2004)., Alla dessa rapporter tyder på att sambandet mellan BR och auxin spelar en viktig roll i regleringen växttillväxt och utveckling.
BR reglerar Auxin-signalering
förutom det ömsesidiga beroendet och samarbetet mellan auxin-och BR-signaler under växtutvecklingen kan BR medla auxin-signalvägen på flera nivåer. BZR1 interagerar med ARF proteiner för att direkt rikta sig mot flera auxin signalering komponenter och gener som är involverade i auxin ämnesomsättning såsom transport-och signalsystem, inklusive AUX/IAA, PINs, TIR1 och ARFs, etc. (Sun et al., 2010)., Det konstaterades att Aux/IAA proteiner som är inblandade i BR svar och iaa7/axr2-1 och iaa17/axr3-3 mutanter visade avvikande BR känslighet och avvikande BR-inducerad genuttryck i ett organ-beroende sätt (Nakamura et al., 2006). Exogent brassinolid (BL) behandling kan inducera uttrycket av auxin-responsiva gener som iaa5, iaa19, iaa17, etc., och uttrycket av ovanstående gener är nedreglerat i BR biosyntetisk mutant de-etioled2 (det2), vilket indikerar att funktionell BR biosyntes delvis krävs för auxinberoende genuttryck (Nakamura et al.,, 2003; Kim et al., 2006). Dessutom, BR auxin påverkar också flödet genom att reglera uttrycket av auxin exportörer som PIN4 och PIN7 (Nakamura et al., 2004). Under växtens gravitropismsvar kan BRs förbättra den polära ackumuleringen av auxin-exportören PIN2 i rotmeristemzonen och därmed påverka omfördelningen av auxin från rotspetsen mot förlängningszonerna och resultera i skillnaden i IAA-nivåer i både övre och nedre sidor av rötter för att inducera växtgravitropism., Under denna process spelar BR-aktiverad ROP2 en viktig roll vid modulering av den funktionella lokaliseringen av PIN2 genom reglering av montering/ommontering av F-aktiner (Li et al., 2005). Ytterligare studier visade att minskad BL perception och/eller koncentration kan inducera CYP79B2, genen som kodar för ett enzym som omvandlar tryptofan till indol-3-acetaldoxim och därmed påverka distributionen (Kim et al., 2007).
dessutom konstaterades det att BR-signalen kunde reglera auxinsignalutgången genom sin negativa regulator GSK3-kinasbin2., Auxin response factor ARF2 identifierades som ett BIN2-interagerande protein i en jäst tvåhybrid skärm och kinasanalys visade BIN2 kunde fosforylat ARF2. Fosforyleringen av ARF2 resulterar i förlust av dess DNA-bindningsförmåga och repressionsaktivitet hos målgenerna (Vert et al., 2008). ARF2 är en bzr1 målgener och dess uttryck reduceras genom BR behandling (Sun et al., 2010)., Dessutom, BIN2 kan phosphorylate ARF7 och ARF19 att undertrycka deras interaktion med AUX/IAAs och därmed öka transkriptionell aktivitet på deras mål gener LATERALA ORGEL GRÄNSER-DOMAIN16 (LBD16) och LBD29 att reglera laterala root organogenes (Cho et al., 2014). Men, BR spelar en mindre roll under denna process och BIN2 är under kontroll av TRACHEARY ELEMENT DIFFERENTIERING HÄMMANDE FAKTOR (TDIF)–TDIF-RECEPTORN (TDR) modul (Cho et al., 2014). Tillsammans kan BR reglera auxin-reponser genom att påverka olika auxin-signaleringskomponenter.,
Auxin reglerar BR-signalering
å andra sidan kan auxin också reglera BR-signalbanan i vissa aspekter. Uttrycket av DVÄRG4, ett avgörande hydroxylas för BR-biosyntes för att kontrollera endogen BR-nivå, är auxinberoende. Auxin behandling kan märkbart stimulera uttryck för DWARF4 och auxin kan hämma bindningen av BZR1 till arrangören av DWARF4. Induktionen av DWARF4 av auxin kräver auxin signaleringsväg men inte BR signaleringsväg (Chung et al., 2011; Yoshimitsu et al., 2011)., CPD katalysera C-3 oxidation av BR aktiverades av BRX, en förmodad transkriptionsfaktor som agerar nedströms av auxin signalering (Mouchel et al., 2006). Ytterligare studier i ris indikerar att exogen auxin kan förbättra transkriptionsuttryck nivåer av BR-receptorgen OsBRI1, vilket tyder på att auxin förbättrar BR-signalering genom reglering av BR-receptorer (Sakamoto et al., 2013). Dessutom har arrangören av osbri1 ett auxre-motiv (upstream auxin-response element) som är inriktat på arf-transkriptionsfaktorer., Dessutom visar mutantstudier att vid mutation av AuxRE avskaffas induktionen av uttryck av osbri1 av auxin och även uttrycket av OsBRI1 regleras i arf-mutant (Sakamoto et al., 2013). Det har rapporterats att OsARF19 binder till arrangören av OsBRI1 och positivt reglerar dess uttryck som sedan aktiverar BR signalering (Zhang et al., 2015). BES1 kan binda till promotorn av SMÅ AUXIN-UPP-RNA 15 (SAUR15) och medla BR tidigt svar gen i Arabidopsis, och denna bindning kan förbättras genom auxin behandling (Walcher och Nemhauser, 2012)., Tillsammans kan auxin också påverka BR-svar och BR-reglerad växttillväxt och utveckling.
avslutande kommentarer och framtida perspektiv
under de senaste nästan fyra decennierna har studier om auxin-BR-interaktioner dragit till sig allt fler forskares intresse. Apparaten av fysiologiska, molekylära, genetiska och biokemiska verktyg har fördjupat vår förståelse för denna fråga., Baserat på tidigare studier är BR och auxin involverade synergistiskt i flera växtutvecklingsprocesser, inklusive: hypokotylförlängning, utveckling av vaskulära buntar, rotutveckling och tropismer etc. Auxin och BR: s ömsesidiga beroende och samarbete är komplicerade och involverar många processer på molekylär nivå genom att dela samma målgener och reglera varandra ömsesidigt på flera nivåer (Figur 1).
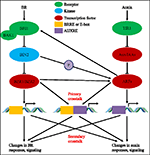
figur 1. Modell av auxin-brassinosteroid (BR) överhörning., I Arabidopsis, uppfattningar av BR och auxin signal är erkända av BRI1 och TIR1 receptorer, respektive. BR binder till den extracellulära domänen av BRI1 och främja det interagerar med co-receptor BAK1 för att bilda en mer aktiv BR receptor-komplex, vilket i sin tur leder till dephosphorylation och inaktivering av BIN2. Inaktivering av BIN2 leda till dephosphorylation av två BR homolog transkriptionsfaktorer BZR1 och BZR2, som flyttar in i kärnan för att aktivera transkription av gener som innehåller BRRE eller E-box i deras promotorregion., BIN2 kan också fosforylatera ARF7 och ARF19 för att undertrycka deras interaktion med AUX/IaaS och därigenom förbättra transkriptionsaktiviteten på deras målgener. TIR1 kvitto auxin signalering och interagera med AUX/IAA proteiner som co-receptorn av auxin. AUX / IAA bryts sedan ned genom ubiquitin-proteasome-vägen, och auxin-transkriptionsregulatorerna auxin response factors (ARFs) frigörs från AUX/IAA-repression och aktiverar transkription av gener med auxin responsive elements (AUXRE) i deras regleringsområde., Vissa ARFs kan också binder till arrangören av BRI1 och positivt reglerar dess uttryck som sedan aktiverar BR signalering. Primär crosstalk sker genom aktivering av gener som innehåller både BRRE / e-box och AUXRE i deras promotorregion, vilket gör att båda signalvägarna direkt reglerar transkription. Sekundär crosstalk sker genom uttryck av gener som är antingen auxin eller BR lyhörd, men de aktiviteter som styr uttryck av gener som reglerar svaret och signalering av andra hormoner.,
Fosforyleringsreglering spelar en avgörande roll i BR-signaleringsvägen, särskilt under perceptionsprocessen, br uppfattas genom BRI1-kinasreceptorn och BAK1-kinaskoreceptorer, och kontrollerar så småningom BR-reglerat genuttryck genom att påverka nedströms transkriptionsfaktorer som BES1/BZR1-aktiviteter (he et al., 2005; Yin et al., 2005; Sun et al., 2010; Tang et al., 2011; Yu et al., 2011). Ubiquitinationsförordningen verkar dock vara nödvändig för auxin-signalering., När auxin binder till TIR1-receptorn, som fungerar som en ubiquitin E3-ligase, den aktiverade TIR1 E3-ligase ubiquitinates AUX/IAA proteiner, leder till nedbrytning av dessa repressors och de-represses ARF transkriptionsfaktorer, och så småningom orsakar auxin reglerade mönster genuttryck förändringar och tillväxt svar (Gray et al., 1999, 2002; Hellmann m.fl., 2003; Quint et al., 2005). Eftersom det har konstaterats att BIN2 kinas, som är kända att fungera i BR signalering, kunde phosphorylate och förbättra verksamheten i ARFs som ARF2 och ARF7 (Vert et al., 2008; Cho m.fl.,, 2014), det ska bli intressant att testa om kinaser som BIN2, som är involverade i BR signalering, kan också interagera med andra auxin signalering komponenter såsom TIR1 receptor eller AUX/IAA repressors och inflytande TIR1 E3-ligase aktivitet eller AUX/IAA protein stabilities. Å andra sidan måste ubiquitinations roll i BR-signalering också åtgärdas, speciellt om TIR1 E3-ligas direkt kan interagera med BR-signaleringskomponenter och reglera deras proteinstabiliteter.,
dessutom, med hjälp av auxin response DR5 och andra auxin reportrar, har det observerats att auxin reglerar växttillväxt och utveckling på ett vävnads-eller cellberoende sätt. De olika transkriptionella utgångarna beroende på cell-och miljökontexten (Clark et al., Och med 2014. Etchells et al., 2016; Lavy et al., 2016)., Om spatiotemporal BR signalering har visat att kontrollera för root tillväxt genom antagonistiska effekten med auxin (Chaiwanon och Wang, 2015), är det fortfarande okänt om vävnaden eller cellulära BR signalering, vilket kan åskådliggöras genom att pBZR1:BZR1-YFP, är också viktigt för att styra andra processer utöver rot-utveckling. Vidare är generering av en detaljerad vävnads – eller källarkarta av auxin-och BR-distributioner för närvarande möjlig med fluorescensaktiverad cellsortering eller lasermikrodissektion i kombination med högupplösande genuttrycksanalys., Detta kommer så småningom leder till adress om auxin crosstalks med BR i en vävnad eller cellulära sätt.
författare bidrag
alla författare var inblandade i skrivandet av denna översyn manuskript. För mer information om vad som utgör författarskap, se våra författarriktlinjer.,
Finansieringen
Detta arbete stöds genom bidrag från National Natural Science Foundation of China (Projekt 31500229 och 31770305), Shandong-Provinsen Natural Science Foundation of Major Basic Research Program (2017C03), Shandong Regionala Medel för att Unga och Framstående Forskare (2014JQ201408).
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
Clouse, S. D. (2011)., Brassinosteroid signaltransduktion: från receptorkinasaktivering till transkriptionella nätverk som reglerar växtutveckling. Växtcell 23, 1219-1230. doi: 10.1105 / TPC.111.084475
PubMed Abstrakt | CrossRef Full Text | Google Scholar
Estelle, M. (2011). Auxin signalering: från syntes till systembiologi; en Ämnessamling från Cold Spring Harbor perspektiv i biologi. Cold Spring Harbor, NEW york: Cold Spring Harbor Laboratory Tryck.
Google Scholar
Guilfoyle, T. J. (2015)., The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response. Plant Cell 27, 33–43. doi: 10.1105/tpc.114.132753
PubMed Abstract | CrossRef Full Text | Google Scholar